Нуклеотиды a c t g. Нуклеотиды. Смотреть что такое "нуклеотиды" в других словарях
- это сложные мономеры, из которых собраны гетерополимерные молекулы. ДНК и РНК. Свободные нуклеотиды участвуют в сигнальных и энергетических процессах жизнедеятельности. ДНК-нуклеотиды и РНК-нуклеотиды имеют общий план строения, но различаются по строению сахара-пентозы. В ДНК-нуклеотидах используется сахар дезоксирибоза, а в РНК-нуклеотидах - рибоза.
Структура нуклеотида
В каждом нуклеотиде можно выделить 3 части:
1. Углевод - это пятичленный сахар-пентоза (рибоза или дезоксирибоза).
2. Фосфорный остаток (фосфат) - это остаток фосфорной кислоты.
3. Азотистое основание - это соединение, в котором много атомов азота. В нуклеиновых кислотах используется всего 5 видов азотистых оснований: Аденин, Тимин, Гуанин, Цитозин, Урацил. В ДНК - 4 вида: Аденин, Тимин, Гуанин, Цитозин. В РНК - тоже 4 вида: Аденин, Урацил, Гуанин, Цитозин, Легко заметить, что в РНК происходит замещение Тимина на Урацил по сравнению с ДНК.
Общая структурная формула пентозы (рибозы или дезоксирибозы), молекулы которой образуют "скелет" нуклеиновых кислот:
Если Х заменить на Н (Х = Н) - то получаются дезоксирибонуклеозиды; если Х заменить на ОН (Х = ОН) - то получаются рибонуклеозиды. Если вместо R подставить азотистое основание (пуриновое или пиримидиновое) - то получится конкретный нуклеотид.
Важно обратить внимание на те положения атомов углерода в пентозе, которые обозначены как 3" и 5". Нумерация атомов углерода начинается от атома кислорода вверху и идёт по часовой стрелке. Последним получается атом углерода (5"), который располагается за пределами пентозного кольца и образует, можно сказать, "хвостик" у пентозы. Так вот, при наращивании цепочки из нуклеотидов фермент может присоединить новый нуклеотид только к углероду 3" и ни к какому другому. Поэтому 5"-конец нуклеотидной цепочки никогда не сможет иметь продолжения, удлинняться может только 3"-конец.



Сравните нуклеотид для РНК с нуклеотидом для ДНК.
Попробуйте узнать, какой это нуклеотид, в таком представлении:

АТФ - свободный нуклеотид
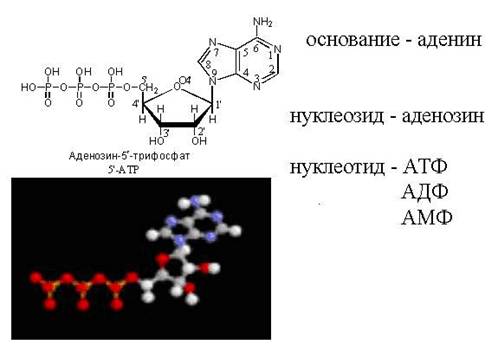
цАМФ - "закольцованная" молекула АТФ
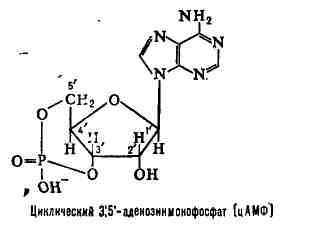
Схема строения нуклеотида
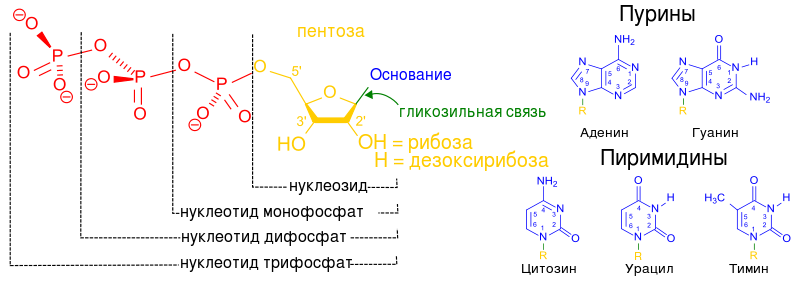
Обратите внимание на то, что активированный нуклеотид, способный наращивать цепочку ДНК или РНК, имеет "трифосфатный хвостик". Именно этим "энергонасыщенным" хвостиком он может присоединиться к уже имеющейся цепочке растущей нуклеиновой кислоты. Фосфатный хвостик сидит на 5-м атоме углерода, так что это положение углерода уже занято фосфатами и предназнено для прикрепления. К чему же его прикрепить? Только к углероду в положении 3". После прикрепления данный нуклеотид сам станет мишенью дла прикрепления следующего нуклеотида. "Принимающая сторона" предоставляет углерод в положении 3", а "прибывающая сторона" цепляется к нему фосфатным хвостиком, находящимся в положении 5". В целом цепочка растёт со стороны 3".
Наращивание нуклеотидной цепочки ДНК
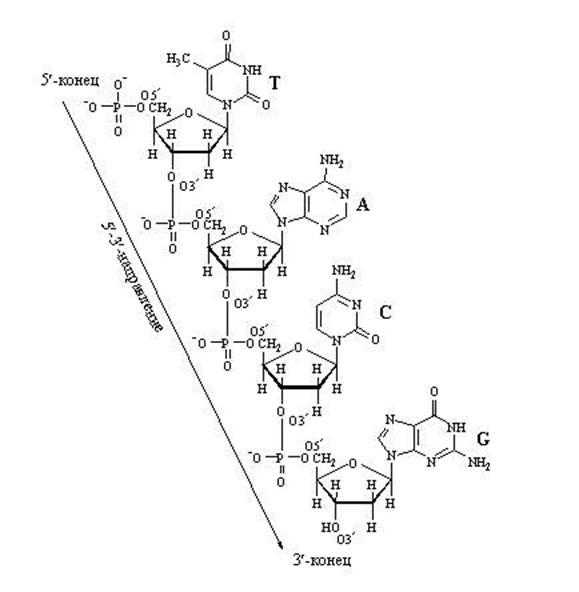
Наращивание цепочки за счёт "продольных" связей между нуклеотидами может идти только в одном направлении: от 5" ⇒ к 3", т.к. новый нуклеотид можно присоединить только к 3"-концу цепочки, но не к 5"-концу.
Пары нуклеотидов, связанные "поперечными" комплементарными связями своих азотистых оснований

Участок двойной спирали ДНК

Найдите признаки антипараллельности двух цепей ДНК.
Найдите пары нуклеотидов с двойными и тройными комплементарными связями.
Лекция № 19
НУКЛЕОЗИДЫ. НУКЛЕОТИДЫ. НУКЛЕИНОВЫЕ КИСЛОТЫ
План
- Нуклеиновые основания.
- Нуклеозиды.
- Нуклеотиды.
- Нуклеотидные коферменты.
- Нуклеиновые кислоты.
Лекция № 19
НУКЛЕОЗИДЫ. НУКЛЕОТИДЫ. НУКЛЕИНОВЫЕ
КИСЛОТЫ
План
- Нуклеиновые основания.
- Нуклеозиды.
- Нуклеотиды.
- Нуклеотидные коферменты.
- Нуклеиновые кислоты.
Нуклеиновые кислоты – присутствующие в
клетках всех живых организмов биополимеры, которые выполняют важнейшие функции
по хранению и передаче генетической информации и участвуют в механизмах ее
реализации в процессе синтеза клеточных белков.
Установление состава нуклеиновых кислот путем их последовательного
гидролитического расщепления позволяет выделить следующие структурные
компоненты.
Рассмотрим структурные компоненты нуклеиновых
кислот в порядке усложнения их строения.
1. Нуклеиновые основания.
Гетероциклические основания, входящие в состав
нуклеиновых кислот (нуклеиновые основания
), — это гидрокси- и
аминопроизводные пиримидина и пурина. Нуклеиновые кислоты содержат три
гетероциклических основания с пиримидиновым циклом (пиримидиновые
основания
) и два — с пуриновым циклом
(пуриновые основания
).
Нуклеиновые основания
имеют тривиальные названия и соответствующие однобуквенные обозначения.

В составе нуклеиновых кислот гетероциклические
основания находятся в термодинамически стабильной оксоформе.
Кроме этих групп нуклеиновых оснований,
называемых основными
, в нуклеиновых кислотах в небольших количествах
встречаются минорные
основания: 6-оксопурин (гипоксантин),
3-N-метилурацил, 1-N-метилгуанин и др.
Нуклеиновые кислоты включают остатки
моносахаридов – D-рибозы и 2-дезокси –D-рибозы. Оба моносахарида присутствуют в
нуклеиновых кислотах в
b
-фуранозной форме.

2. Нуклеозиды.
Нуклеозиды – это N-гликозиды, образованные нуклеиновыми основаниями и рибозой
или дезоксирибозой.
Между аномерным атомом углерода моносахарида и атомом азота в положении 1
пиримидинового цикла или атомом азота в положении 9 пуринового цикла образуется b
-гликозидная
связь.

В зависимости от природы моносахаридного остатка
нуклеозиды делят на рибонуклеозиды
(содержат остаток рибозы) и дезоксирибонуклеозиды
(содержат остаток дезоксирибозы). Названия
нуклеозидов строят на основе тривиальных названий нуклеиновых оснований,
добавляя окончание –идин
для производных пиримидина и -озин
для
производных пурина. К названиям дезоксирибонуклеозидов добавляют приставку дезокси-.
Исключение составляет нуклеозид, образованный тимином и
дезоксирибозой, к которому приставка дезокси-
не добавляется, так как
тимин образует нуклеозиды с рибозой лишь в очень редких случаях.

Для обозначения нуклеозидов используются
однобуквенные обозначения, входящих в их состав нуклеиновых оснований. К
обозначениям дезоксирибонуклеозидов (за исключением тимидина) добавляется буква
”д”.
Наряду с представленными на схеме основными
нуклеозидами в составе нуклеиновых кислот встречаются минорные нуклеозиды,
содержащие модифицированные нуклеиновые основания (см. выше).
В природе нуклеозиды встречаются также в
свободном состоянии, преимущественно в виде нуклеозидных антибиотиков, которые
проявляют противоопухолевую активность. Нуклеозиды-антибиотики имеют некоторые
отличия от обычных нуклеозидов в строении либо углеводной части, либо
гетероциклического основания, что позволяет им выступать в качестве
антиметаболитов, чем и объясняется их антибиотическая активность.
Как N-гликозиды, нуклеозиды устойчивы к действию
щелочей, но расщепляются под действием кислот с образованием свободного
моносахарида и нуклеинового основания. Пуриновые нуклеозиды гидролизуются
значительно легче пиримидиновых.
3. Нуклеотиды
Нуклеотиды – это эфиры нуклеозидов и фосфорной
кислоты (нуклеозидфосфаты). Сложноэфирную связь с фосфорной кислотой образует ОН
группа в положении 5
/ или
3
/ моносахарида. В зависимости от
природы моносахаридного остатка нуклеотиды делят на рибонуклеотиды
(структурные элементы РНК) и дезоксирибонуклеотиды
(структурные элементы
ДНК). Названия нуклеотидов включают название нуклеозида с указанием положения в
нем остатка фосфорной кислоты. Сокращенные обзначения нуклеозидов содержат
обозначение нуклеозида, остатка моно-, ди- или трифосфорной кислоты, для
3
/ -производных указывается также
положение фосфатной группы.

Нуклеотиды являются мономерными звеньями, из
которых построены полимерные цепи нуклеиновых кислот. Некоторые нуклеотиды
выполняют роль коферментов и участвуют в обмене веществ.
4. Нуклеотидные
коферменты
Коферменты
– это органические соединения
небелковой природы, которые необходимы для осуществления каталитического
действия ферментов. Коферменты относятся к разным классам органических
соединений. Важную группу коферментов составляют нуклеозидполифосфаты
.
Аденозинфосфаты –
производные
аденозина, содержащие остатки моно-, ди- и трифосфорных кислот. Особое место
занимают аденозин-5 / -моно-, ди- и
трифосфаты — АМФ, АДФ и АТФ — макроэргические
вещества, которые обладают
большими запасами свободной энергии в подвижной форме. Молекула АТФ содержит
макроэргические связи Р-О, которые легко расщепляются в результате гидролиза.
Выделяющаяся при этом свободная энергия обеспечивает протекание сопряженных с
гидролизом АТФ термодинамически невыгодных анаболических процессов, например,
биосинтез белка.

Кофермент А
.
Молекула этого
кофермента состоит из трех структурных компонентов: пантотеновой кислоты,
2-аминоэтантиола и АДФ.

Кофермент А участвует в процессах
ферментативного ацилирования, активируя карбоновые кислоты путем превращения их
в реакционноспособные сложные эфиры тиолов.
Никотинамидадениндинуклеотидные коферменты.
Никотинамидадениндинуклеотид (НАД +)
и его фосфат (НАДФ
+ ) содержат в своем составе катион пиридиния в виде
никотинамидного фрагмента. Пиридиниевый катион в составе этих коферментов
способен обратимо присоединять гидрид-анион с образованием восстановленной формы
кофермента — НАД
Н.

Таким образом никотинамидадениндинуклеотидные
коферменты участвуют в окислительно-восстановительных процессах, связанных с
переносом гидрид-аниона, например, окислении спиртовых групп в альдегидные
(превращение ретинола в ретиналь), восстановительном аминировании кетокислот,
восстановлении кетокислот в гидроксикислоты. В ходе этих процессов субстрат
теряет (окисление) или присоединяет (восстановление) два атома водорода в виде
Н
+ и Н
— . Кофермент служит при этом акцептором
(НАД
+ ) или донором
(НАД .
Н) гидрид-иона. Все процессы с
участием коферментов являются стереоселективными. Так, при восстановлении
пировиноградной кислоты образуется исключительно L-молочная кислота.

5. Нуклеиновые кислоты.
Первичная структура
нуклеиновых кислот представляет собой линейную полимерную цепь, построенную
из мономеров – нуклеотидов, которые связаны между собой
3 / -5 / -фосфодиэфирными
связями. Полинуклеотидная цепь имеет 5′-конец и 3′- конец. На 5′-конце находится
остаток фосфорной кислоты, а на 3′- конце — свободная гидроксильная группа.
Нуклеотидную цепь принято записывать, начиная с 5′-конца.

В зависимости от природы моносахаридных остатков
в нуклеотиде различают дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые
кислоты (РНК). ДНК и РНК различаются также по природе входящих в их состав
нуклеиновых оснований: урацил входит только в состав РНК, тимин – только в
состав ДНК.
Вторичная структура
ДНК
представляет собой комплекс двух полинуклеотидных цепей, закрученных вправо
вокруг общей оси так, что углевод-фосфатные цепи находятся снаружи, а
нуклеиновые основания направлены внутрь (двойная спираль Уотсона-Крика
).
Шаг спирали — 3.4 нм, на 1 виток приходится 10 пар нуклеотидов. Полинуклеотидные
цепи антипараллельны,т.е.
напротив 3′-конца одной цепи находится 5′-конец другой цепи. Две цепи ДНК
неодинаковы по своему составу, но они комплементарны
. Это выражается в
том, что напротив аденина (А) в одной цепи всегда находится тимин (Т) в другой
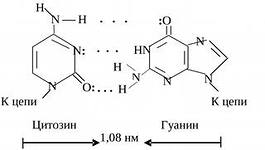
цепи, а напротив гуанина (Г) всегда находится цитозин (Ц). Комплементарное
спаривание А с Т и Г с Ц осуществляется за счет водородных связей. Между А и Т
образуется две водородные связи, между Г и Ц – три.

Комплементарность цепей ДНК составляет
химическую основу важнейшей функции ДНК – хранения и передачи генетической
информации.
Типы РНК.
Известны три основных
вида клеточных РНК: транспортные РНК (тРНК), матричные РНК (мРНК) и рибосомные
РНК (рРНК). Они различаются по месторасположению в клетке, составу и размерами,
а также функциями. РНК состоят, как правило, из одной полинуклеотидной цепи,
которая в пространстве складывается таким образом, что ее отдельные участки
становятся комплементарными друг другу (”слипаются”) и образуют короткие
двуспиральные участки молекулы, в то время как другие участки остаются
однотяжевыми.
Матричные РНК
выполняют функцию матрицы
белкового синтеза в рибосомах.
Рибосомные РНК
выполняют роль структурных
компонентов рибосом.
Транспортные РНК
участвуют в
транспортировке
a
-аминокислот из цитоплазмы в рибосомы и в переводе информации нуклеотидной
последовательности мРНК в последовательность аминокислот в белках.
Механизм передачи генетической информации.
Генетическая информация закодированиа в нуклеотидной последовательности
ДНК. Механизм передачи этой информации включает три основных этапа.
Первый этап – репликация
–копирование
материнской ДНК с образованием двух дочерних молекул ДНК, нуклеотидная
последовательность которых комплементарна последовательности материнской ДНК и
однозначно определяется ею. Репликация осуществляется путем синтеза новой
молекулы ДНК на материнской, которая играет роль матрицы. Двойная спираль
материнской ДНК раскручивается и на каждой из двух цепей происходит синтез новой
(дочерней) цепи ДНК с учетом принципа комплементарности. Процесс осуществляется
под действием фермента ДНК-полимеразы. Таким образом из одной материнской ДНК
образуются две дочерних, каждая из которых содержит в своем составе одну
материнскую и одну вновь синтезированную полинуклеотидную цепь.
Второй этап – транскрипция
– процесс, в
ходе которого часть генетической информации переписывается с ДНК в форме мРНК.
Матричная РНК синтезируется на участке деспирализованной цепи ДНК как на матрице
под действием фермента РНК-полимеразы. В полинуклеотидной цепи мРНК
рибонуклеотиды, несущие
определенные
нуклеиновые основания, выстраиваются в последовательности, определяемой
комплементарными взаимодействиями с нуклеиновыми основаниями цепи ДНК. При этом адениновому
основанию в ДНК будет соответствовать урациловое
основание в РНК. Генетическая информация о синтезе белка закодирована в ДНК с
помощью триплетного
кода. Одна аминокислота кодируется
последовательностью из трех нуклеотидов, которую называют кодоном
.
Участок ДНК, кодирующий одну полипептидную цепь, называется геном
.
Каждому кодону ДНК соответствует комплементарный кодон в мРНК. В целом молекула
мРНК комплементарна определенной части цепи ДНК – гену.
Процессы репликации и транскрипции происходят в
ядре клетки. Синтез белка осуществляется в рибосомах. Синтезированная мРНК
мигрирует из ядра в цитоплазму к рибосомам, перенося генетическую информацию к
месту синтеза белка.
Третий этап – трансляция
– процесс
реализации генетической информации, которую несет мРНК в виде последовательности
нуклеотидов в последовательность аминокислот в синтезируемом белке.
a
-Аминокислоты, необходимые для
синтеза белка транспортируются к рибосомам посредством тРНК, с которыми они
связываются путем ацилирования 3
/ -ОН группы на конце цепи тРНК.

тРНК имеет антикодоновую ветвь, содержащую
тринуклеотид — антикодон
, который соответствует переносимой ею
аминокислоте. На рибосоме тРНК прикрепляются антикодоновыми участками к
соответствующим кодонам мРНК. Специфичность стыковки кодона и антикодона
обеспечивается их комплементарностью. Между сближенными аминокислотами
образуется пептидная связь. Таким образом реализуется строго определенная
последовательность соединения аминокислот в белки, закодированная в
генах.
Учебное пособие предназначено для студентов направления «Биология» всех профилей подготовки, всех форм обучения для теоретической подготовки к занятиям, зачетам и экзаменам. Пособие охватывает основные разделы структурной биохимии: строение, физико-химические свойства и функции основных классов биологических макромолекул. Большое внимание уделено ряду прикладных аспектов биохимии.
Нуклеотиды и нуклеиновые кислоты
Структура нуклеотидов и азотистых оснований
Нуклеотиды принимают участие во множестве биохимических процессов, а также являются мономерами нуклеиновых кислот. Нуклеиновые кислоты обеспечивают все генетические процессы. Каждый нуклеотид состоит из трех типов химических молекул:
Азотистое основание;
Моносахарид;
1-3 остатка фосфорной кислоты.
В отличие от моносахаридов, нуклеотиды как мономеры являются сложно устроенными молекулами, состоящими из структур, относящихся к разным классам химических веществ, поэтому необходимо рассмотреть свойства и структуру этих компонентов по отдельности.
Азотистые основания
Азотистые основания относятся к гетероциклическим соединениям. В состав гетероцикла помимо атомов углерода входят атомы азота. Все азотистые основания, входящие в нуклеотиды относят к двум классам азотистых оснований: пуриновые и пиримидиновые. Пуриновые основания это производные пурина – гетероцикла, состоящего из двух циклов, один пятичленный, второй – шести, нумерация осуществляется так, как показано на рисунке. Пиримидиновые основания являются производными пиримидина и состоят из одного шестичленного цикла, нумерация также указана на рисунке (Рисунок 31). Главные пиримидиновые основания и у прокариот, и у эукариот – это цитозин, тимин и урацил. Из пуриновых оснований чаще всего встречаются аденин и гуанин. Два других – ксантин и гипоксантин – являются интермедиатами в процессах их метаболизма. У человека в роли конечного продукта катаболизма пуринов выступает окисленное пуриновое основание – мочевая кислота . Помимо пяти названных выше главных оснований известны и менее широко представленные минорные основания. Некоторые из них присутствуют только в нуклеиновых кислотах бактерий и вирусов, но многие также найдены в составе про- и эукариотических ДНК и транспортных и рибосомных РНК. Так, и бактериальная ДНК, и ДНК человека содержат значительные количества 5-метилцитозина; в бактериофагах обнаружен 5-гидроксиметилцитозин. Необычные основания выявлены в матричной РНК – N 6 -метиладенин, N 6 , N 6 -диметиладенин и N 7 -Meтилгуанин. У бактерий также обнаружен модифицированный урацил с присоединенной по N 3 -положению (α-амино, α-карбокси) -пропильной группой. Функции этих замещенных пуринов и пиримидинов до конца не выяснены, однако они могут образовывать неканонические связи между основаниями (это будет рассмотрено ниже), обеспечивая образование вторичных и третичных структур нуклеиновых кислот.
Рисунок 31. Структура азотистых оснований
В клетках растений выявлена серия пуриновых оснований с метильными заместителями. Многие из них фармакологически активны. В качестве примера можно привести кофейные зерна, содержащие кофеин (1,3, 7-триметилксантин), чайный лист, содержащий теофиллин (1, 3-диметил-ксантин), и какао-бобы, в состав которых входит теобромин (3, 7-диметилксантин).
изомерия и Физико-химические свойства пуриновых и пиримидиновых оснований
Молекула азотистого основания образует систему чередующихся одинарных и двойных связей (систему сопряженных двойных связей). Такая организация образует жесткую молекулу, без возможности конформационных переходов. В результате нельзя говорить об изменении конформации азотистых оснований.
Для азотистых оснований выявлен только один тип изомерии кето-енольный переход или таутомерия.
Таутомерия
Благодаря феномену кето-енольной таутомерии нуклеотиды могут существовать либо в лактимной, либо в лактамной формах, причем в физиологических условиях лактамная форма превалирует у гуанина и тимина (Рисунок 32). Важность этого обстоятельства станет ясна при обсуждении процессов спаривания оснований.
Рисунок 32. Таутомерия нуклеотидов
Растворимость
При нейтральном рН наименьшей растворимостью обладает гуанин. Следующим в этом ряду стоит ксантин. Мочевая кислота в форме уратов сравнительно неплохо растворяется при нейтральном рН, но очень плохо растворима в жидкостях с более низкими значениями рН, таких, как моча. Гуанин в моче человека в норме отсутствует, а ксантин и мочевая кислота являются ее обычными компонентами. Последние два пурина часто входят в состав камней мочевого тракта.
Поглощение света
За счет системы сопряженных двойных связей все азотистые основания поглощают в ультрафиолетовой части спектра. Спектр поглощения – график распределения оптической плотности в зависимости от длины волны. Для каждого азотистого основания свой спектр поглощения, по нему можно различить растворы различных азотистых оснований или соединений в состав которых входит азотистое основание (нуклеотиды), но максимум поглощения у всех совпадает при длине волны 260 нм. Это позволяет легко и быстро определять концентрацию как азотистых оснований, так нуклеотидов и нуклеиновых кислот. Спектр поглощения также зависит от рН раствора (Рисунок 33).
Рисунок 33. Спектры поглощения различных азотистых оснований
Функции азотистых оснований
Азотистые основания практически не встречаются в свободном состоянии. Исключение составляют некоторые алкалоиды и мочевая кислота.
Азотистые основания выполняют следующие функции:
Входят в состав нуклеотидов;
Часть алкалоидов – азотистые основания, например, кофеин в кофе или теофелин в чае;
Промежуточные продукты обмена азотистых оснований и нуклеотидов;
Мочевая кислота – причина мочекаменной болезни;
В виде мочевой кислоты выводится азот у некоторых организмов.
Нуклеотиды и нуклеозиды
Молекулы нуклеозидов построены из пуринового или пиримидинового основания, к которому (β-связью присоединен углевод (обычно D-рибоза или 2-дезоксирибоза) в N 9 или N 1 ‒положении соответственно. Таким образом, адениновый рибонуклеозид (аденозин) состоит из аденина и D-рибозы, присоединенной в положении N 9 ; гуанозин – из гуанина и D-рибозы в положении N 9 ; цитидин – из цитозина и рибозы в положении N 1 ; уридин – из урацила и рибозы в положении N 1 . Таким образом в пуриновых нуклеозидах (нуклеотидах) азотистое основание и сахар связаны 1-9 β гликозидной связью, а в пиримидинах – 1-1 β гликозидной связью.
В состав 2́-дезоксирибонуклеозидов входят пуриновые или пиримидиновые основания и 2́-дезоксирибоза, присоединенная по тем же атомам N 1 и N 9 . Присоединение рибозы или 2́-дезоксирибозы к кольцевой структуре основания происходит за счет относительно кислотолабильной N-гликозидной связи (Рисунок 34).
Нуклеотиды – это производные нуклеозидов, фосфорилированные по одной или более гидроксильным группам остатка рибозы (или дезоксирибозы). Так, аденозинмонофосфат (AMФ или аденилат) построен из аденина, рибозы и фосфата. 2́-дезоксиаденозинмонофосфат (дAMФ или дезоксиаденилат) представляет собой молекулу, состоящую из аденина, 2́-дезоксирибозы и фосфата. Обычно к урацилу присоединена рибоза, к тимину – 2́-дезоксирибоза. Поэтому тимидиловая кислота (ТМФ) состоит из тимина, 2́-дезоксирибозы и фосфата. Кроме вышеперечисленных форм нуклеотидов обнаружены и нуклеотиды необычной структуры. Так, в молекуле тРНК выявлен нуклеотид, в котором рибоза присоединяется к урацилу в пятом положении, т. е. не азот-углеродной связью, а углерод-углеродной. Продукт этого необычного присоединения назван псевдоуридином (ψ). Молекулы тРНК содержат и другую необычную нуклеотидную структуру – тимин, соединенный с рибозомонофосфатом. Этот нуклеотид образуется уже после синтеза молекулы тРНК путем метилирования остатка УMФ S-аденозилметионином. Псевдоуридиловая кислота (ψМФ) тоже образуется в результате перегруппировки УMФ после синтеза тРНК.
Рисунок 34. Структура пуриновых и пиримидиновых нуклеозидов и нуклеотидов
Номенклатура, физико-химические свойства и функции нуклеозидов и нуклеотидов
Положение фосфатной группы в молекуле нуклеотида указывается цифрой. Например, аденозин с фосфатной группой, присоединенной к 3-му углероду рибозы, должен быть обозначен как 3́-монофосфат. Штрих после цифры ставят для того, чтобы отличить номер углерода в пуриновом или пиримидиновом основании от положения этого атома в остатке дезоксирибозы. При нумерации атомов углерода основания штрих не ставится. Нуклеотид 2́-дезоксиаденозин с фосфатным остатком при углероде-5 молекулы сахара обозначается как 2́-дезоксиаденозин-5́-монофосфат. Нуклеозиды, содержащие аденин, гуанин, цитозин, тимин и урацил, принято обозначать буквами A, Г, Ц, Т и У соответственно. Наличие буквы d (или д) перед сокращением обозначает, что углеводным компонентом нуклеозида является 2́-дезоксирибоза. Гуанозин, содержащий 2́-дезоксирибозу, может быть обозначен дГ (дезоксигуанозин), а соответствующий ему монофосфат с фосфатной группой, присоединенной к третьему атому углерода дезоксирибозы, – дГ-3́-МФ. Как правило, в тех случаях, когда фосфат присоединен к углероду-5 рибозы или дезоксирибозы, символ 5́ опускается. Так, гуанозин 5́-монофосфат принято обозначать ГМФ, а 5́-монофосфат 2́-дезоксигуанозина сокращают как дГМФ. Если к углеводному остатку нуклеозида присоединены 2 или 3 остатка фосфорной кислоты используются аббревиатуры ДФ (дифосфат) и ТФ (трифосфат). Таким образом, аденозин + трифосфат с тремя фосфатными группами в 5́-положении углевода будет обозначаться АТФ. Поскольку в молекулах нуклеотидов фосфаты находятся в виде ангидридов фосфорной кислоты, т. е. в состоянии с низкой энтропией, их называют макроэргами (обладающими большим запасом потенциальной энергии). При гидролизе 1 моля АТФ до AДФ высвобождается 7,3 кКал потенциальной энергии.
Рисунок 35. Структура цАМФ
Физико-химические свойства нуклеотидов
Так как в состав нуклеотидов входят азотистые основания, то такие свойства как таутомерия и способность поглощать в ультрафиолетовой части спектра также характерны и для нуклеотидов, причем спектры поглощения азотистых оснований и содержащих эти основания нуклеотидов сходны. Наличие сахара и остатков фосфорной кислоты делает их более гидрофильными чем азотистые основания. Все нуклеотиды являются кислотами, так как содержат остатки фосфорной кислоты.
Функции природных нуклеотидов
Нуклеотиды являются мономерами нуклеиновых кислот (РНК, ДНК). В состав ДНК входят дезоксирибонуклеотидфосфаты – производные аденина, тимина, гуанина и цитозина. Также некоторые молекулы гуанина и цитозина в составе ДНК метилированы, то есть содержат метильную группу. Как основные мономеры в состав РНК входят рибонуклеотидфосфаты – производные аденина, урацила, гуанина и цитозина. Также в состав РНК входят нуклеотиды, содержащие различные минорные азотистые основания, например ксантин, гипоксантин, дигидроуридин и др.
Нуклеотиды являются мономерами коферментов (НАД, НАДФ, ФАД, ко-энзим А, метионин-аденозин). В составе коферементов они участвуют в ферментативных реакциях. Более подробно эта функция будет рассмотрена ниже.
Энергетическая (АТФ) . АТФ выполняет функцию основного внутриклеточного переносчика свободной энергии. Концентрация наиболее распространенного свободного нуклеотида в клетках млекопитающих – АТФ – составляет около 1 ммоль/л.
Сигнальная (цГМФ, цАМФ) (Рисунок 35). Циклический AMФ (3́-, 5́-аденозинмонофосфат, цАМФ) – медиатор различных внеклеточных сигналов в клетках животных – образуется из АТФ в результате реакции, катализируемой аденилатциклазой. Активность аденилатциклазы регулируется комплексом взаимодействий, многие из которых инициируются через рецепторы гормонов. Внутриклеточная концентрация цАМФ (около 1 мкмоль/л) на 3 порядка ниже концентрации ATФ. Циклический цГМФ (3́-, 5́-гуанозинмонофосфат, цГМФ) служит внутриклеточным проводником внеклеточных сигналов. В некоторых случаях цГМФ выступает в роли антагониста цАМФ. цГМФ образуется из ГТФ под действием гуанилатциклазы – фермента, имеющего много общего с аденилатциклазой. Гуанилатциклаза, как и аденилатциклаза, регулируется различными эффекторами, в том числе и гормонами. Как и цАМФ, цГМФ гидролизуется фосфодиэстеразой до соответствующего 5́-монофосфата.
Регуляторная (ГТФ) . Активность группы белков (G-белков), выполняющих в основном регуляторную функцию, зависит от того: какой нуклеотид они связывают. В неактивной форме эти белки связывают ГДФ, при активации белка происходит замена ГДФ на ГТФ. При выполнении своей функции белок гидролизует ГТФ до ГДФ и фосфата, выделившаяся, энергия затрачивается на функционирование белка.
Активация при метаболизме липидов и моносахаридов (УТФ, СТФ) . Производные урациловых нуклеотидов участвуют в качестве активирующих агентов в реакциях метаболизма гексоз и полимеризации углеводов, в частности при биосинтезе крахмала и олигосахаридных фрагментов гликопротеинов и протеогликанов. Субстратами в этих реакциях являются уридин-дифосфатсахара. Например, уридиндифосфатглюкоза служит предшественником гликогена. Также превращение глюкозы в галактозу, глюкуроновую кислоту или другие производные моносахаридов происходит в виде коньюгата с УДФ. СТР необходим для биосинтеза некоторых фосфоглицеридов в тканях животных. Реакции с участием церамида и ЦДФ-холина приводят к образованию сфингомиелина и других замещенных сфингозинов.
Участие в дезактивации различных спиртов и фенолов (УДФ-глюкуроновая кислота). Уридиндифосфатглюкуроновая кислота – выполняет функцию «активного» глюкуронида в реакциях конъюгирования, например, при образовании глюкуронида билирубина.
Нуклеотиды в составе коферментов
Коферменты – это низкомолекулярные соединения связанные с ферментами (см раздел «Ферменты») непосредственно участвующие в в биохимической реакции, другими словами это еще один субстрат, не выходящий в окружающую среду.
Коферменты подразделяют на две группы:
переносчики протонов и электронов, эти коферменты участвуют в окислительно-восстановительных реакциях;
переносчики всех остальных групп кроме протонов и электронов, эти коферменты участвуют в трансферазных реакциях.
Более подробно механизмы упомянутых реакций можно рассмотреть в главе «Ферменты».
Некоторые коферменты содержат в своем составе нуклеотиды. Они также делятся на эти же две группы.
Коферменты переносчики протонов и электронов
Эти коферменты участвуют в окислительно-восстановительных реакциях, где аденозин выполняет только структурную функцию, в реакцию вступают нуклеотиды, содержащие другие типы оснований, выделяют два типа таких коферментов: никотиновые и флавиновые. Они отличаются не только по активной группировке, но и по типу реакций, которые они осуществляют.
Никотиновые коферменты
Рисунок 36. Никотиновые коферменты. А-структура NAD, Б-структура NADP, В-механизм активности никотиновой кислоты, Г-механизм работы никотиновых коферментов
Никотинамидадениндинуклеотид (NAD +) – главный акцептор электронов при окислении топливных молекул. Реакционноспособная часть NAD + – его никотинамидное кольцо. При окислении субстрата никотинамидное кольцо NAD + присоединяет ион водорода и два электрона, которые являются эквивалентами гидрид-иона. Восстановленная форма этого переносчика – NADH. В ходе этого дегидрирования один атом водорода субстрата прямо переносится на NAD + , тогда как второй переходит в растворитель. Оба электрона, теряемые субстратом, переносятся на никотинамидное кольцо. Роль донора электронов в большинстве процессов восстановительного биосинтеза (пластического обмена); выполняет восстановленная форма никотин амидадениндинуклеотидфосфата (NADPH). NADPH отличается от NAD наличием фосфата, связанного эфирной связью с 2́-гидроксильной группой аденозина. Окисленная форма NADPH обозначается как NADP + . NADPH переносит электроны таким же образом, как NADH. Однако, NADPH используется почти исключительно в процессах восстановительного биосинтеза, тогда как NADH используется преимущественно для генерирования АТР. Дополнительная фосфатная группа NADPH – это участок, ответственный за осуществление целевого предназначения молекулы, состоящего в распознавании ферментами.
Флавиновые коферменты
Первый флавиновый кофермент (флавинмононуклеотид FMN) был выделен А. Сент-Дьёрдьи из сердечной мышцы в 1932 г., Р. Г. Варбург и В. Христиан тогда же получили из дрожжей первый флавопротеид, содержащий FMN в качестве кофермента. Второй важнейший флавиновый кофермент – флавинадениндинуклеотид (FAD) выделен ими же как кофактор оксидазы D-аминокислот в 1938 году. За счет окислительно-восстановительного превращения флавинового кольца флавиновые коферменты осуществляют окислительно-восстановительные реакции в составе многих важнейших ферментных систем: оксидаз (в частности, оксидаз D- и L-аминокислот, моноаминооксидазы, регулирующей уровень катехоламинов в крови) и дегидрогеназ (часто с участием никотинамидадениндинуклеотида и убихинонов).
Рисунок 37. Флавиновые коферменты. А-структура FAD, Б-механизм активности никотиновой кислоты, В-механизм работы флавиновых коферментов
Второй основной переносчик электронов при окислении топливных молекул – флавинадениндинуклеотид. Сокращения, используемые для обозначения окисленной и восстановленной форм этого переносчика – соответственно FAD и FADH 2 . Реакционноспособная часть FAD – это его изоаллоксазиновое кольцо. FAD, подобно NAD + , присоединяет два электрона. Однако FAD в отличие от NAD + присоединяет оба теряемых субстратом атома водорода.
Конец ознакомительного фрагмента.
Нуклеозиды, в свою очередь, являются N-гликозидами , содержащими гетероциклический фрагмент, связанный через атом азота с C-1 атомом остатка сахара .
В природе наиболее распространены нуклеотиды, являющиеся β -N-гликозидами пуринов или пиримидинов и пентоз - D-рибозы или D-2-дезоксирибозы. В зависимости от структуры пентозы различают рибонуклеотиды и дезоксирибонуклеотиды , которые являются мономерами молекул сложных биологических полимеров (полинуклеотидов) - соответственно РНК или ДНК .
Фосфатный остаток в нуклеотидах обычно образует сложноэфирную связь с 2′-, 3′- или 5′-гидроксильными группами рибонуклеозидов, в случае 2′-дезоксинуклеозидов этерифицируются 3′- или 5′-гидроксильные группы.
Большинство нуклеотидов являются моноэфирами ортофосфорной кислоты, однако известны и диэфиры нуклеотидов, в которых этерифицированы два гидроксильных остатка - например, циклические нуклеотиды циклоаденин- и циклогуанин монофосфаты (цАМФ и цГМФ). Наряду с нуклеотидами - эфирами ортофосфорной кислоты (монофосфатами) в природе также распространены и моно- и диэфиры пирофосфорной кислоты (дифосфаты, например, аденозиндифосфат) и моноэфиры триполифосфорной кислоты (трифосфаты, например, аденозинтрифосфат).
Номенклатура
| Код | Означает | Комплементарная пара |
|---|---|---|
| A | T в ДНК ; U в РНК |
|
| C | G | |
| G | C | |
| или |
T в ДНК ; U в РНК |
A |
| M | A или C |
K |
| R | A или G |
Y |
| W | A или T |
W |
| S | C или G |
S |
| Y | C или T |
R |
| K | G или T |
M |
| V | A или C или G |
B |
| H | A или C или T |
D |
| D | A или G или T |
H |
| B | C или G или T |
V |
| X или N |
A или C или G или T (U) |
любой |
Соединения, состоящие из двух нуклеотидовых молекул, называются динуклеотидами , из трёх - тринуклеотидами , из небольшого числа - олигонуклеотидами , а из многих - полинуклеотидами , или нуклеиновыми кислотами .
Названия нуклеотидов представляют собой аббревиатуры в виде стандартных трёх- или четырёхбуквенных кодов.
Если аббревиатура начинается со строчной буквы «д» (англ. d ), значит подразумевается дезоксирибонуклеотид ; отсутствие буквы «д» означает рибонуклеотид . Если аббревиатура начинается со строчной буквы «ц» (англ. c ), значит речь идёт о циклической форме нуклеотида (например, цАМФ).
Первая прописная буква аббревиатуры указывает на конкретное азотистое основание или группу возможных нуклеиновых оснований, вторая буква - на количество остатков фосфорной кислоты в структуре (М - моно-, Д - ди-, Т - три-), а третья прописная буква - всегда буква Ф («-фосфат »; англ. P ).
Латинские и русские коды для нуклеиновых оснований:
- T - Т: Тимин (5-метилурацил) , встречается у бактериофагов в ДНК , занимает место урацила в РНК ;
- U - У: Урацил , встречается в РНК , занимает место тимина в ДНК .
Общепринятые буквенные коды для обозначения нуклеотидных оснований соответствуют номенклатуре, принятой Международным союзом теоретической и прикладной химии (International Union of Pure and Applied Chemistry, сокращённо - англ. IUPAC , ИЮПАК) и Международным союзом биохимии и молекулярной биологии (International Union of Biochemistry and Molecular Biology (англ.) русск. , сокращённо - англ. IUBMB ). Если при секвенировании последовательности ДНК или РНК возникает сомнение в точности определения того или иного нуклеотида, помимо пяти основных (A, C, T, G, U), используют другие буквы латинского алфавита в зависимости от того, какие наиболее вероятные нуклеотиды могут находиться в данной позиции последовательности. Эти же дополнительные буквы используют для обозначения вырожденных (не совпадающих у разных гомологичных последовательностей) позиций, например при записи последовательности праймеров для ПЦР.
Длину секвенированных участков ДНК (гена , сайта , хромосомы) или всего генома указывают в парах нуклеотидов (пн), или парах оснований (англ. base pairs , сокращённо bp ), подразумевая под этим элементарную единицу двухцепочечной молекулы нуклеиновой кислоты, сложенную из двух спаренных комплементарных оснований.
Биологическая роль
1. Универсальный источник энергии (АТФ и его аналоги).
2. Являются активаторами и переносчиками мономеров в клетке(УДФ-глюкоза)
6. Являются мономерами в составе нуклеиновых кислот, связанные 3′-5′-фосфодиэфирными связями .
История
В домолекулярной генетике для обозначения наименьшего элемента в структуре ДНК , который может быть подвержен спонтанной или индуцированной мутации , применялся особый термин рекон . В настоящее время показано, что таким наименьшим элементом является один нуклеотид (или одно азотистое основание в составе нуклеотида), поэтому данный термин более не употребляется. Для определения понятия единица мутации применялся термин мутон . В настоящее время показано, что фенотипически мутация может проявиться даже при замене одного нуклеотида (или азотистого основания в составе нуклеотида), таким образом, термин мутон соответствует одному нуклеотиду.
Напишите отзыв о статье "Нуклеотиды"
Примечания
См. также
Ссылки
- Нуклеотиды - статья из Большой советской энциклопедии .
- Буквенные коды нуклеотидных оснований согласно номенклатуре ИЮПАК и IUBMB (англ.) русск.
- в интернет-журнала
|
||||||||||||||||||||||||||||||||||
Отрывок, характеризующий Нуклеотиды
Казака призвали, расспросили; казачьи командиры хотели воспользоваться этим случаем, чтобы отбить лошадей, но один из начальников, знакомый с высшими чинами армии, сообщил этот факт штабному генералу. В последнее время в штабе армии положение было в высшей степени натянутое. Ермолов, за несколько дней перед этим, придя к Бенигсену, умолял его употребить свое влияние на главнокомандующего, для того чтобы сделано было наступление.– Ежели бы я не знал вас, я подумал бы, что вы не хотите того, о чем вы просите. Стоит мне посоветовать одно, чтобы светлейший наверное сделал противоположное, – отвечал Бенигсен.
Известие казаков, подтвержденное посланными разъездами, доказало окончательную зрелость события. Натянутая струна соскочила, и зашипели часы, и заиграли куранты. Несмотря на всю свою мнимую власть, на свой ум, опытность, знание людей, Кутузов, приняв во внимание записку Бенигсена, посылавшего лично донесения государю, выражаемое всеми генералами одно и то же желание, предполагаемое им желание государя и сведение казаков, уже не мог удержать неизбежного движения и отдал приказание на то, что он считал бесполезным и вредным, – благословил совершившийся факт.
Записка, поданная Бенигсеном о необходимости наступления, и сведения казаков о незакрытом левом фланге французов были только последние признаки необходимости отдать приказание о наступлении, и наступление было назначено на 5 е октября.
4 го октября утром Кутузов подписал диспозицию. Толь прочел ее Ермолову, предлагая ему заняться дальнейшими распоряжениями.
– Хорошо, хорошо, мне теперь некогда, – сказал Ермолов и вышел из избы. Диспозиция, составленная Толем, была очень хорошая. Так же, как и в аустерлицкой диспозиции, было написано, хотя и не по немецки:
«Die erste Colonne marschiert [Первая колонна идет (нем.) ] туда то и туда то, die zweite Colonne marschiert [вторая колонна идет (нем.) ] туда то и туда то» и т. д. И все эти колонны на бумаге приходили в назначенное время в свое место и уничтожали неприятеля. Все было, как и во всех диспозициях, прекрасно придумано, и, как и по всем диспозициям, ни одна колонна не пришла в свое время и на свое место.
Когда диспозиция была готова в должном количестве экземпляров, был призван офицер и послан к Ермолову, чтобы передать ему бумаги для исполнения. Молодой кавалергардский офицер, ординарец Кутузова, довольный важностью данного ему поручения, отправился на квартиру Ермолова.
– Уехали, – отвечал денщик Ермолова. Кавалергардский офицер пошел к генералу, у которого часто бывал Ермолов.
– Нет, и генерала нет.
Кавалергардский офицер, сев верхом, поехал к другому.
– Нет, уехали.
«Как бы мне не отвечать за промедление! Вот досада!» – думал офицер. Он объездил весь лагерь. Кто говорил, что видели, как Ермолов проехал с другими генералами куда то, кто говорил, что он, верно, опять дома. Офицер, не обедая, искал до шести часов вечера. Нигде Ермолова не было и никто не знал, где он был. Офицер наскоро перекусил у товарища и поехал опять в авангард к Милорадовичу. Милорадовича не было тоже дома, но тут ему сказали, что Милорадович на балу у генерала Кикина, что, должно быть, и Ермолов там.
– Да где же это?
– А вон, в Ечкине, – сказал казачий офицер, указывая на далекий помещичий дом.
– Да как же там, за цепью?
– Выслали два полка наших в цепь, там нынче такой кутеж идет, беда! Две музыки, три хора песенников.
Офицер поехал за цепь к Ечкину. Издалека еще, подъезжая к дому, он услыхал дружные, веселые звуки плясовой солдатской песни.
«Во олузя а ах… во олузях!..» – с присвистом и с торбаном слышалось ему, изредка заглушаемое криком голосов. Офицеру и весело стало на душе от этих звуков, но вместе с тем и страшно за то, что он виноват, так долго не передав важного, порученного ему приказания. Был уже девятый час. Он слез с лошади и вошел на крыльцо и в переднюю большого, сохранившегося в целости помещичьего дома, находившегося между русских и французов. В буфетной и в передней суетились лакеи с винами и яствами. Под окнами стояли песенники. Офицера ввели в дверь, и он увидал вдруг всех вместе важнейших генералов армии, в том числе и большую, заметную фигуру Ермолова. Все генералы были в расстегнутых сюртуках, с красными, оживленными лицами и громко смеялись, стоя полукругом. В середине залы красивый невысокий генерал с красным лицом бойко и ловко выделывал трепака.
– Ха, ха, ха! Ай да Николай Иванович! ха, ха, ха!..
Офицер чувствовал, что, входя в эту минуту с важным приказанием, он делается вдвойне виноват, и он хотел подождать; но один из генералов увидал его и, узнав, зачем он, сказал Ермолову. Ермолов с нахмуренным лицом вышел к офицеру и, выслушав, взял от него бумагу, ничего не сказав ему.
– Ты думаешь, это нечаянно он уехал? – сказал в этот вечер штабный товарищ кавалергардскому офицеру про Ермолова. – Это штуки, это все нарочно. Коновницына подкатить. Посмотри, завтра каша какая будет!
На другой день, рано утром, дряхлый Кутузов встал, помолился богу, оделся и с неприятным сознанием того, что он должен руководить сражением, которого он не одобрял, сел в коляску и выехал из Леташевки, в пяти верстах позади Тарутина, к тому месту, где должны были быть собраны наступающие колонны. Кутузов ехал, засыпая и просыпаясь и прислушиваясь, нет ли справа выстрелов, не начиналось ли дело? Но все еще было тихо. Только начинался рассвет сырого и пасмурного осеннего дня. Подъезжая к Тарутину, Кутузов заметил кавалеристов, ведших на водопой лошадей через дорогу, по которой ехала коляска. Кутузов присмотрелся к ним, остановил коляску и спросил, какого полка? Кавалеристы были из той колонны, которая должна была быть уже далеко впереди в засаде. «Ошибка, может быть», – подумал старый главнокомандующий. Но, проехав еще дальше, Кутузов увидал пехотные полки, ружья в козлах, солдат за кашей и с дровами, в подштанниках. Позвали офицера. Офицер доложил, что никакого приказания о выступлении не было.
– Как не бы… – начал Кутузов, но тотчас же замолчал и приказал позвать к себе старшего офицера. Вылезши из коляски, опустив голову и тяжело дыша, молча ожидая, ходил он взад и вперед. Когда явился потребованный офицер генерального штаба Эйхен, Кутузов побагровел не оттого, что этот офицер был виною ошибки, но оттого, что он был достойный предмет для выражения гнева. И, трясясь, задыхаясь, старый человек, придя в то состояние бешенства, в которое он в состоянии был приходить, когда валялся по земле от гнева, он напустился на Эйхена, угрожая руками, крича и ругаясь площадными словами. Другой подвернувшийся, капитан Брозин, ни в чем не виноватый, потерпел ту же участь.
Все живое на планете состоит из многочисленных клеток. Они поддерживают упорядоченность своей организации с помощью генетической информации, содержащейся в ядре, которая сохраняется, передается и реализуется высокомолекулярными сложными соединениями — нуклеиновыми кислотами. Кислоты эти, в свою очередь, состоят из мономерных звеньев – нуклеотидов.
Вконтакте
Роль нуклеиновых кислот переоценить невозможно. Нормальная жизнедеятельность организма определяется стабильностью их структуры. Если в строении происходят любые отклонения, меняется количество либо последовательность — это обязательно приводит к изменениям в клеточной организации. Изменяется активность физиологических процессов и жизнедеятельность клеток .
Понятие нуклеотида
 Как и белки, нуклеиновые кислоты необходимы для жизни
. Это генетический материал всех живых организмов, включая вирусы.
Как и белки, нуклеиновые кислоты необходимы для жизни
. Это генетический материал всех живых организмов, включая вирусы.
Выяснение структуры одного из двух типов нуклеиновых кислот ДНК позволило понять, каким образом в живых организмах хранится информация, необходимая для регулирования жизнедеятельности и как она передается потомству. Нуклеотид представляет собой мономерную единицу, образующую соединения более сложные — нуклеиновые кислоты. Без них невозможно хранение , воспроизведение и передача генетической информации. Свободные нуклеотиды – главные компоненты, которые участвуют в энергетических и в сигнальных процессах. Они поддерживают нормальную жизнедеятельность отдельных клеток и организма в целом.Из них строятся длинные молекулы — полинуклеотиды.Чтобы разобраться со структурой полинуклеотида следует понять строение нуклеотидов.
Что такое нуклеотид? Молекулы ДНК собраны из мелких мономерных соединений. Другими словами, нуклеотид — это органическое сложное соединение, представляющее собой составную часть нуклеиновых кислот и других биологических соединений, необходимых для жизнедеятельности клетки.
Состав и основные свойства нуклеотидов
 В состав молекулы нуклеотида (мононуклеотида) в определенной последовательности входят три химических соединения:
В состав молекулы нуклеотида (мононуклеотида) в определенной последовательности входят три химических соединения:
- Пентоза или пятиугольный сахар:
- дезоксирибоза. Эти нуклеотиды называют дезоксирибонуклеотидами. Они входят в состав ДНК;
- рибоза. Нуклеотиды входят в состав РНК и называются рибонуклеотидами.
2. Азотистая пиримидиновая или пуриновая основа, связанная с углеродным атомом сахара. Это соединение называют нуклеозидом
3. Фосфатная группа, состоящая из остатков фосфорной кислоты (в количестве от одного до трех). Присоединяется к углероду сахара эфирными связями, образующими молекулу нуклеотида.
Свойствами нуклеотидов являются:
- участие в метаболизме и других физиологических процессах, которые протекают в клетке;
- осуществление контроля над репродукцией и ростом;
- хранение информации о наследуемых признаках и о структуре белка.
Нуклеиновые кислоты
 Сахар в нуклеиновых кислотах представлен пентозой. В РНК пятиуглеродный сахар называется рибозой, в ДНК - дезоксирибозой. В каждой молекуле пентозы пять атомов углерода, из которых четыре образуют кольцо с атомом кислорода, а пятый атом входит в группу НО-СН2.
Сахар в нуклеиновых кислотах представлен пентозой. В РНК пятиуглеродный сахар называется рибозой, в ДНК - дезоксирибозой. В каждой молекуле пентозы пять атомов углерода, из которых четыре образуют кольцо с атомом кислорода, а пятый атом входит в группу НО-СН2.
В молекуле положение атома углерода обозначается цифрой со штрихом (например:1C´, 3C´, 5C´). Так как у вех процессов считывания с молекулы нуклеиновой кислоты наследственной информации имеется строгая направленность, нумерация углеродных атомов и их расположение служат указателем верного направления.
С первым углеродным атомом 1C´ в молекуле сахара соединяется азотистое основание.
К третьему и пятому углеродным атомам по гидроксильной группе (3C´, 5C´) присоединяется остаток фосфорной кислоты, который определяет химическую принадлежность к группе кислот ДНК и РНК.
Состав азотистых оснований
 Виды нуклеотидов по азотистому основанию ДНК:
Виды нуклеотидов по азотистому основанию ДНК:
Первые два класса — пурины:
- аденин (А);
- гуанин (Г).
Два последние относятся к классу пиримидинов:
- тимин (Т);
- цитозин (Ц).
Пуриновые соединения по молекулярной массе тяжелее пиримидиновых.
Нуклеотиды РНК по азотистому соединению представлены:
- гуанином;
- аденином;
- урацитолом;
- цитозином.
Так же, как тимин, урацил является пиримидиновым основанием. Нередко в научной литературе азотистые основания обозначаются латинскими буквами (A, T, C, G, U).
Пиримидины, а именно тимин, цитозин, урацил представлены шестичленным кольцом, состоящим из двух атомов азота и четырех атомов углерода, последовательно пронумерованных, от 1 до 6.
Пурины (гуанин и аднин) состоят из имидазола и пиримидина. В молекулах пуриновых оснований четыре атома азота и пять атомов углерода. У каждого атома имеется свой номер от 1 дот 9.
Результатом соединений азотистых остатков с остатками пентозы является нуклеозид. Нуклеотид – это соединение фосфатной группы с нуклеозидом.
Образование фосфодиэфирных связей
 Следует разобраться в вопросе о том, как нуклеотиды соединяются в полипептидную цепь, сколько их участвует в процессе,образуя молекулу нуклеиновой кислоты за счет фосфодиэфирных связей.
Следует разобраться в вопросе о том, как нуклеотиды соединяются в полипептидную цепь, сколько их участвует в процессе,образуя молекулу нуклеиновой кислоты за счет фосфодиэфирных связей.
При взаимодействии двух нуклеотидов образуется динуклеотид. Новое соединение образуется путем конденсации, когда возникает фосфодиэфирная связь между гидроксигруппой пентозы одного мономера и фосфатным остатком другого.
Синтезом полинуклеотида является многочисленное повторение этой реакции. Сборка полинуклеотидов представляет сложный процесс, обеспечивающей рост цепи с одного конца.
 Молекулы ДНК, как и молекулы белка, имеют первичную, вторичную структуры и третичную. Первичную структуру в цепи ДНК определяет последовательность нуклеотидов. В основе вторичной структуры лежит формирование водородных связей. При синтезе двойной спирали ДНК
имеется определенная закономерность и последовательность: тимин одной цепи соответствует аденину другой; цитозин – гуанину, и наоборот. Соединения нуклеидов создают прочную связь цепей, с равным между ними расстоянием.
Молекулы ДНК, как и молекулы белка, имеют первичную, вторичную структуры и третичную. Первичную структуру в цепи ДНК определяет последовательность нуклеотидов. В основе вторичной структуры лежит формирование водородных связей. При синтезе двойной спирали ДНК
имеется определенная закономерность и последовательность: тимин одной цепи соответствует аденину другой; цитозин – гуанину, и наоборот. Соединения нуклеидов создают прочную связь цепей, с равным между ними расстоянием.
Зная последовательность нуклеотидов одной цепи ДНК можно по принципу дополнения или комплементарности достроить вторую.
Третичная структура ДНК образовывается путем трехмерных сложных соединений. Это делает молекулу более компактной, чтобы она могла свободно разместиться в небольшом объеме клетки. длина кишечной палочки ДНК более 1 мм, в то время как длина самой клетки менее 5 мкм.
Количество пиримидиновых оснований равняется всегда числу пуриновых. Расстояние между нуклеотидами равняется 0,34 нм. Это постоянная величина, как и молекулярная масса.
Функции и свойства ДНК
 Основные функции ДНК:
Основные функции ДНК:
- сохраняет наследственную информацию;
- передача (удвоение/репликация);
- транскрипция, реализация;
- ауторепродукция ДНК. Функционирование репликона.
Процесс самовоспроизведения молекулы нуклеиновой кислоты сопровождается передачей от клетки к клетке копий генетической информаций. Для его осуществления необходимы набор специфических ферментов. В этом процессе полуконсервативного типа образуется репликативная вилка.
Репликон представляет собой единицу репликационного процесса участка генома, подконтрольного одной точке инициации репликации. Как правило, геном прокариот -это репликон. Репликация от точки инициации идет в обе стороны, иногда с различной скоростью.
Молекула РНК – структура
 РНК является одной полинуклеотидной цепочкой, которая образуется через ковалентные связи между фосфатным остатком и пентозой. Она короче ДНК, имеет другую последовательность и различается по видовому составу азотистых соединений. Пиримидиновое основание тимина в РНК
заменяется урацилом.
РНК является одной полинуклеотидной цепочкой, которая образуется через ковалентные связи между фосфатным остатком и пентозой. Она короче ДНК, имеет другую последовательность и различается по видовому составу азотистых соединений. Пиримидиновое основание тимина в РНК
заменяется урацилом.
РНК может быть трех видов, в зависимости от тех функций, которые выполняются в организме:
- информационная (иРНК) — очень разнообразная по нуклеотидному составу. Она является своего рода матрицей для синтеза белковой молекулы, переносит генетическую информацию к рибосомам от ДНК;
- транспортная (тРНК) в среднем состоит из 75-95 нуклеотидов. Она переносит необходимую аминокислоту в рибосоме к месту синтеза полипептида. У каждого вида тРНК и есть своя, присущая только ему последовательность нуклеотидов или мономеров;
- рибосомальная (рРНК) обычно одержит от 3000 до 5000 нуклеотидов. Рибосом является необходимым структурным ом компонент участвующим в важнейшем процессе, происходящем в клетке – биосинтезе белка.
Роль нуклеотида в организме
В клетке нуклеотиды выполняют важные функции:
- являются биорегуляторами;
- используются как структурные блоки для нуклеиновых кислот;
- входят в состав главного источника энергии в клетке — АТФ;
- участвуют во многочисленных обменных процессах в клетках;
- являются переносчиками восстановительных эквивалентов в клетках (ФАД, НАДФ+; НАД+; ФМН);
- могут рассматриваться как вестники регулярного внеклеточного синтеза (цГМФ, цАМФ).
Свободные нуклеотиды – главные компоненты, которые участвуют в энергетических и в сигнальных процессах. Они поддерживают нормальную жизнедеятельность отдельных клеток и организма в целом.