Формирование мембранного потенциала покоя. Происхождение потенциала покоя Уменьшение разности потенциалов в мембране клетки называется
Потенциал покоя
Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин:
1) неодинакового распределения ионов по обе стороны мембраны;
2) избирательной проницаемости мембраны для ионов.
В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ.
За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концент-рации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил:
1) силы диффузии;
2) силы электростатического взаимодействия. Значение электрохимического равновесия:
1) поддержание ионной асимметрии;
2) поддержание величины мембранного потенциала на постоянном уровне.
В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентра-ционно-электрохимическим.
Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ.
Потенциал действия
Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны.
При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный.
Компоненты потенциала действия:
1) локальный ответ;
2) высоковольтный пиковый потенциал (спайк);
3) следовые колебания.
Ионы Na путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это то количество милливольт, на которое должен снизиться мембранный потенциал, чтобы возник лавинообразный ход ионов Na в клетку.
Высоковольтный пиковый потенциал (спайк).
Пик потенциала действия является постоянным компонентом потенциала действия. Он состоит из двух фаз:
1) восходящей части – фазы деполяризации;
2) нисходящей части – фазы реполяризации.
Лавинообразное поступление ионов Na в клетку приводит к изменению потенциала на клеточной мембране. Чем больше ионов Na войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется активационных ворот. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны. Движение ионов Na внутрь клетки продолжается до момента электрохимического равновесия по иону Na Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов Na и от степени проницаемости мембраны к ионам Na. Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. При достижении электрохимического равновесия по ионам Na происходит инактивация активационных ворот, снижается проницаемость к ионам Na и возрастает проницаемость к ионам K. Полного восстановления мембранного потенциала не происходит.
В процессе восстановительных реакций на клеточной мембране регистрируются следовые потенциалы – положительный и отрицательный.
В данной теме будут рассмотрены два катиона – натрий (Na) и калий (К). Говоря об анионах, примем во внимание то, что определенное количество анионов находится у наружной и внутренней стороны мембраны клетки.
Форма клетки зависит от того, к какой ткани она принадлежит. По своей форме клетки могут быть:
· цилиндрические и кубические (клетки кожи);
· дисковидные (эритроциты);
· шаровидные (яйцеклетки);
· веретеновидные (гладкомышечные);
· звездчатые и пирамидные (нервные клетки);
· не имеющие постоянной формы – амебовидные (лейкоциты).
Клетка обладает рядом свойств: она питается, растет, размножается, восстанавливается, адаптируется к окружающей её среде, обменивается энергией и веществами с окружающей средой, выполняет присущие ей функции (в зависимости от того, к какой ткани принадлежит данная клетка). Кроме того, клетка обладает возбудимостью.
Возбудимость – это способность клетки в ответ на раздражения переходить из состояния покоя к состоянию активности.
Раздражения могут поступать из внешней среды или возникать внутри клетки. Раздражителями, вызывающими возбуждение, могут быть: электрические, химические, механические, температурные и иные стимулы.
Клетка может находиться в двух основных состояниях – в покое и в возбуждении. Покой и возбуждение клетки иначе называют – мембранный потенциал покоя и мембранный потенциал действия.
Когда клетка не испытывает ни каких раздражений, она находится в состоянии покоя. Покой клетки иначе называется мембранным потенциалом покоя (МПП).
В состоянии покоя внутренняя поверхность ее мембраны заряжена отрицательно, а наружная – положительно. Это объясняется тем, что внутри клетки находится много анионов и мало катионов, а за клеткой наоборот, преобладают катионы.
Поскольку в клетке присутствуют электрические заряды, то создаваемое ими электричество можно измерить. Величина мембранного потенциала покоя равна: - 70 мВ, (минус 70, поскольку внутри клетки отрицательный заряд). Данная величина условна, так как в каждой клетке может быть своя величина потенциала покоя.
В состоянии покоя поры мембраны открыты для ионов калия и закрыты для ионов натрия. Это означает, что ионы калия могут легко проникать в клетку и выходить из нее. Ионы натрия не могут поступать в клетку, поскольку для них закрыты поры мембраны. Но небольшое число ионов натрия проникает в клетку, потому что притягиваются большим количеством анионов, расположенных на внутренней поверхности мембраны (разноименные заряды притягиваются). Такое перемещение ионов является пассивным, поскольку не требует затрат энергии.
Для нормальной жизнедеятельности клетки величина ее МПП должна оставаться на постоянном уровне. Однако, перемещение ионов натрия и калия через мембрану вызывает колебания данной величины, что может привести к уменьшению или увеличению значения: - 70 мВ.
Для того, чтобы величина МПП оставалась относительно постоянной начинает действовать так называемый натрий – калиевый насос. Его функция заключается в том, что он ионы натрия убирает из клетки, а ионы калия нагнетает в клетку. Именно определенное соотношение ионов натрия и калия в клетке и за клеткой создает нужную величину МПП. Работа насоса является активным механизмом, поскольку для нее требуется энергия.
Источником энергии в клетке является АТФ. АТФ дает энергию только при расщеплении на более простую кислоту - АДФ, при обязательном участии в реакции фермента АТФ-азы:
АТФ + фермент АТФ-аза АДФ + энергия
В покое на наружной стороне плазматической мембраны располагается тонкий слой положительных зарядов, а на внутренней стороне – отрицательных. Разность между ними называется мембранным потенциалом покоя. Если считать наружный заряд равным нулю, то разность зарядов между наружной и внутренней поверхностями у большинства нейронов оказывается близкой к -65 мВ, хотя она и может у отдельных клеток варьировать от -40 до -80 мВ.
Возникновение этой разности зарядов обусловлено неодинаковым распределением ионов калия, натрия и хлора внутри клетки и снаружи её, а также большей проницаемостью покоящейся клеточной мембраны лишь для ионов калия.
У возбудимых клеток мембранный потенциал покоя (МПП) способен сильно изменяться и эта способность является основой для возникновения электрических сигналов. Уменьшение мембранного потенциала покоя, например, с -65 до -60 мВ, называется деполяризацией , а увеличение, например, с -65 до -70 мВ, – гиперполяризацией .
Если деполяризация достигнет некоторого критического уровня, например -55 мВ, то проницаемость мембраны для ионов натрия на короткое время становится максимальной, они устремляются в клетку и в связи с этим трансмембранная разность потенциалов стремительно уменьшается до 0, а затем приобретает положительное значение. Это обстоятельство приводит к закрытию натриевых каналов и стремительному выходу из клетки ионов калия через предназначенные только для них каналы: в результате восстанавливается первоначальная величина мембранного потенциала покоя. Эти быстро происходящие изменения мембранного потенциала покоя называются потенциалом действия. Потенциал действия является приводящимся электрическим сигналом, он быстро распространяется по мембране аксона до самого его окончания, причём нигде не меняет свою амплитуду.
Кроме потенциалов действия в нервной клетке, вследствие изменения её мембранной проницаемости, могут возникать местные или локальные сигналы: рецепторный потенциал и постсинаптический потенциал . Их амплитуда значительно меньше, чем у потенциала действия, кроме того, она существенно уменьшается при распространении сигнала. По этой причине местные потенциалы и не могут распространяться по мембране далеко от места своего возникновения.
Работой натрий-калиевого насоса в клетке создаётся высокая концентрация ионов калия, а в клеточной мембране для этих ионов есть открытые каналы. Выходящие из клетки по концентрационному градиенту ионы калия увеличивают количество положительных зарядов на наружной поверхности мембраны. В клетке много крупномолекулярных органических анионов и потому изнутри мембрана оказывается заряженной отрицательно. Все остальные ионы могут проходить через покоящуюся мембрану в очень небольшом количестве, их каналы, в основном, закрыты. Следовательно, потенциал покоя обязан своим происхождением, главным образом, току ионов калия из клетки .
| |
Электрические сигналы: входной, объединённый, проводящийся и выходной
 Нейроны вступают в контакты с определёнными клетками-мишенями, причём цитоплазма контактирующих клеток не соединяется и между ними всегда сохраняется синаптическая щель.
Нейроны вступают в контакты с определёнными клетками-мишенями, причём цитоплазма контактирующих клеток не соединяется и между ними всегда сохраняется синаптическая щель.
Современный вариант нейронной теории связывает определённые части нервной клетки с характером возникающих в них электрических сигналов. В типичном нейроне есть четыре определяемые морфологически области: дендриты, сома, аксон и пресинаптическое окончание аксона. При возбуждении нейрона в нём последовательно появляется четыре разновидности электрических сигналов: входной, объединённый, проводящийся и выходной (рис. 3.3). Каждый из этих сигналов возникает только в определённой морфологической области.
Входными сигналами являются либо рецепторный , либо постсинаптический потенциал . Рецепторный потенциал образуется в окончаниях чувствительного нейрона, когда на них действует определённый стимул: растяжение, давление, свет, химическое вещество и т.п. Действие стимула вызывает открытие определённых ионных каналов мембраны, а последующий ток ионов через эти каналы изменяет первоначальное значение мембранного потенциала покоя; в большинстве случаев происходит деполяризация. Эта деполяризация и является рецепторным потенциалом, её амплитуда пропорциональна силе действующего стимула.
Рецепторный потенциал может распространяться от места действия стимула вдоль мембраны на относительно небольшое расстояние – амплитуда рецепторного потенциала уменьшается по мере удаления от места действия стимула, а затем деполяризующий сдвиг и вовсе исчезнет.
Вторая разновидность входного сигнала – постсинаптический потенциал . Он образуется на постсинаптической клетке после того, как возбуждённая пресинаптическая клетка отправит для неё нейромедиатор. Добравшись путём диффузии до постсинаптической клетки, медиатор присоединяется к специфическим белкам-рецепторам её мембраны, что вызывает открытие ионных каналов. Возникший в связи с этим ток ионов через постсинаптическую мембрану изменяет первоначальное значение мембранного потенциала покоя – этот сдвиг и является постсинаптическим потенциалом.
В одних синапсах такой сдвиг представляет собой деполяризацию и, если она достигнет критического уровня, то постсинаптический нейрон возбуждается. В других синапсах возникает противоположный по направленности сдвиг: постсинаптическая мембрана гиперполяризуется: величина мембранного потенциала становится больше и уменьшить её до критического уровня деполяризации становится труднее. Такую клетку трудно возбудить, она заторможена. Таким образом, деполяризующий постсинаптический потенциал является возбуждающим , а гиперполяризующий – тормозным . Соответственно этому и сами синапсы подразделяются на возбуждающие (вызывающие деполяризацию) и тормозные (вызывающие гиперполяризацию).
Вне зависимости от того, что происходит на постсинаптической мембране: деполяризация или гиперполяризация, величина постсинаптических потенциалов всегда пропорциональна количеству подействовавших молекул медиатора, но обычно их амплитуда невелика. Так же, как и рецепторный потенциал, они распространяются вдоль мембраны на очень небольшое расстояние, т.е. тоже относятся к местным потенциалам.
Таким образом, входные сигналы представлены двумя разновидностями местных потенциалов, рецепторным и постсинаптическим, а возникают эти потенциалы в строго определённых областях нейрона: либо в чувствительных окончаниях, либо в синапсах. Чувствительные окончания принадлежат сенсорным нейронам, где рецепторный потенциал возникает под действием внешних раздражителей. Для интернейронов, а также для эфферентных нейронов входным сигналом может быть только постсинаптический потенциал.
Объединённый сигнал может возникнуть только в таком участке мембраны, где достаточно много ионных каналов для натрия. В этом отношении идеальным объектом является аксонный холмик – место отхождения аксона от тела клетки, поскольку именно здесь самая высокая во всей мембране плотность каналов для натрия. Такие каналы являются потенциалзависимыми, т.е. открываются лишь тогда, когда исходное значение потенциала покоя достигнет критического уровня. Типичное для среднестатистического нейрона значение потенциала покоя составляет приблизительно -65 мВ, а критический уровень деполяризации соответствует примерно -55 мВ. Стало быть, если удастся деполяризовать мембрану аксонного холмика с -65 мВ до -55 мВ, то там возникнет потенциал действия.
Деполяризовать мембрану способны входные сигналы, т.е. либо постсинаптические потенциалы, либо рецепторные. В случае рецепторных потенциалов местом возникновения объединённого сигнала является ближайший к чувствительным окончаниям перехват Ранвье, где наиболее вероятна деполяризация до критического уровня. Каждый чувствительный нейрон имеет множество окончаний, являющихся ветвями одного отростка. И, если в каждом из этих окончаний при действии стимула возникает очень небольшой по амплитуде рецепторный потенциал и распространяется к перехвату Ранвье с уменьшением амплитуды, то он является лишь малой частью общего деполяризующего сдвига. От каждого чувствительного окончания в одно и то же время перемещаются к ближайшему перехвату Ранвье эти небольшие рецепторные потенциалы, а в области перехвата все они суммируются. Если общая сумма деполяризующего сдвига будет достаточной, то в перехвате возникнет потенциал действия.
Постсинаптические потенциалы, возникающие на дендритах, так же невелики, как и рецепторные потенциалы и так же уменьшаются при распространении от синапса до аксонного холмика, где может возникнуть потенциал действия. Кроме того, на пути распространения постсинаптических потенциалов по телу клетки могут оказаться тормозные гиперполяризующие синапсы и потому возможность деполяризации мембраны аксонного холмика на 10 мВ кажется маловероятной. Тем не менее, этот результат регулярно достигается в результате суммации множества небольших постсинаптических потенциалов, возникающих одновременно в многочисленных синапсах, образованных дендритами нейрона с окончаниями аксонов пресинаптических клеток.
Таким образом, объединённый сигнал возникает, как правило, вследствие суммации одновременно образовавшихся многочисленных местных потенциалов. Такая суммация происходит в том месте, где особенно много потенциалзависимых каналов и поэтому легче достигается критический уровень деполяризации. В случае интеграции постсинаптических потенциалов таким местом является аксонный холмик, а суммация рецепторных потенциалов происходит в ближайшем от чувствительных окончаний перехвате Ранвье (или близко расположенным к ним участком немиелинизированного аксона). Область возникновения объединённого сигнала называется интегративной или триггерной.
Накопление небольших деполяризующих сдвигов молниеносно трансформируется в интегративной зоне в потенциал действия, который является максимальным электрическим потенциалом клетки и возникает по принципу "всё или ничего". Это правило надо понимать так, что деполяризация ниже критического уровня не приносит никакого результата, а при достижении этого уровня всегда, независимо от силы стимулов, обнаруживается максимальный ответ: третьего не дано.
Проведение потенциала действия . Амплитуда входных сигналов пропорциональна силе подействовавшего стимула или количеству выделившегося в синапсе нейромедиатора – такие сигналы называют градуальными . Их длительность определяется длительностью стимула или присутствия медиатора в синаптической щели. Амплитуда и длительность потенциала действия от этих факторов не зависят: оба этих параметра всецело определяются свойствами самой клетки. Стало быть, любая комбинация входных сигналов, любой вариант суммации, при единственном условии деполяризации мембраны до критического значения, вызывает один и тот же стандартный образец потенциала действия в триггерной зоне. Он всегда имеет максимальную для данной клетки амплитуду и примерно одинаковую длительность, сколько бы раз ни повторялись вызывающие его условия.
Возникнув в интегративной зоне, потенциал действия быстро распространяется по мембране аксона. Это происходит благодаря появлению локального электрического тока. Поскольку деполяризованный участок мембраны оказывается иначе заряженным, чем соседствующий с ним, между полярно заряженными участками мембраны возникает электрический ток. Под действием этого локального тока деполяризуется до критического уровня соседний участок, что вызывает появление потенциала действия и в нём. В случае миелинизированного аксона таким соседним участком мембраны является ближайший к триггерной зоне перехват Ранвье, затем следующий, и потенциал действия начинает "перепрыгивать" от одного перехвата к другому со скоростью, достигающей 100 м/с.
Разные нейроны могут многим отличаться друг от друга, но возникающие в них потенциалы действия различить очень трудно, чаще невозможно. Это в высшей степени стереотипный сигнал у самых разных клеток: сенсорных, интернейронов, моторных. Эта стереотипия свидетельствует о том, что сам потенциал действия не содержит никаких сведений о природе породившего его стимула. О силе стимула свидетельствует частота возникающих потенциалов действия, а определением природы стимула занимаются специфические рецепторы и хорошо упорядоченные межнейронные связи.
Таким образом, возникший в триггерной зоне потенциал действия быстро распространяется по ходу аксона к его окончанию. Это передвижение связано с образованием локальных электрических токов, под влиянием которых потенциал действия как бы заново возникает в соседнем участке аксона. Параметры потенциала действия при проведении по аксону нисколько не меняются, что позволяет передавать информацию без искажений. Если аксоны нескольких нейронов оказываются в общем пучке волокон, то по каждому из них возбуждение распространяется изолированно.
Выходной сигнал адресуется другой клетке или одновременно нескольким клеткам и в подавляющем большинстве случаев представляет собой выделение химического посредника – медиатора. В пресинаптических окончаниях аксона заранее запасённый медиатор хранится в синаптических пузырьках, которые накапливаются в специальных участках – активных зонах. Когда потенциал действия добирается до пресинаптического окончания, содержимое синаптических пузырьков путём экзоцитоза опорожняется в синаптическую щель.
Химическими посредниками передачи информации могут служить разные вещества: небольшие молекулы, как, например, ацетилхолин или глутамат, либо достаточно крупные молекулы пептидов – все они специально синтезируются в нейроне для передачи сигнала. Попав в синаптическую щель, медиатор диффундирует к постсинаптической мембране и присоединяется к её рецепторам. В результате связи рецепторов с медиатором изменяется ионный ток через каналы постсинаптической мембраны, а это приводит к изменению значения потенциала покоя постсинаптической клетки, т.е. в ней возникает входной сигнал – в данном случае постсинаптический потенциал.
Таким образом, почти в каждом нейроне, независимо от его величины, формы и занимаемой в цепи нейронов позиции, можно обнаружить четыре функциональные области: локальную рецептивную зону, интегративную, зону проведения сигнала и выходную или секреторную зону (рис. 3.3).
Установлено, что наиболее важными ионами, определяющими мембранные потенциалы клеток, являются неорганические ионы К + , Na + , СГ, а также в ряде случаев Са 2 + . Хорошо известно, что концентрации этих ионов в цитоплазме и в межклеточной жидкости различаются в десятки раз.
Из табл. 11.1 видно, что концентрация ионов К + внутри клетки в 40-60 раз выше, чем в межклеточной жидкости, тогда как для Na + и СГ распределение концентраций противоположное. Неравномерное распределение концентраций этих ионов по обе стороны мембраны обеспечивается как их различной проницаемостью, так и сильным электрическим полем мембраны, которое определяется ее потенциалом покоя.
Действительно, в состоянии покоя суммарный поток ионов через мембрану равен нулю, и тогда из уравнения Не- рнста - Планка следует, что
Таким образом, в покое градиенты концентрации - и
электрического потенциала -- на мембране направлены
противоположно друг другу и поэтому в покоящейся клетке высокая и постоянная разность концентраций основных ионов обеспечивает поддержание на мембране клетки электрического напряжения, которое и называют равновесным мембранным потенциалом.
В свою очередь возникающий на мембране потенциал покоя препятствует выходу ионов из клетки К + и чрезмерному входу в нее СГ, поддерживая тем самым их концентрационные градиенты на мембране.
Полное выражение для мембранного потенциала, учитывающее потоки диффузии этих трех видов ионов, было получено Гольдманом, Ходжкиным и Катцем:
где Р к, P Na , Р С1 - проницаемость мембраны для соответствующих ионов.
Уравнение (11.3) с высокой точностью определяет мембранные потенциалы покоя различных клеток. Из него следует, что для мембранного потенциала покоя важны не абсолютные величины проницаемостей мембраны для различных ионов, а их отношения, так как, разделив обе части дроби под знаком логарифма, например, на Р к, мы перейдем к относительным проницаемостям ионов.
В тех случаях, когда проницаемость одного из этих ионов значительно больше, чем других, уравнение (11.3) переходит в уравнение Нернста (11.1) для этого иона.
Из табл. 11.1 видно, что мембранный потенциал покоя клеток близок к потенциалу Нернста для ионов К + и СВ, но значительно отличается от него по Na + . Это свидетельствует
0 том, что в покое мембрана хорошо проницаема для ионов К + и СГ, тогда как для ионов Na + ее проницаемость очень низка.
Несмотря на то что равновесный потенциал Нернста для СГ наиболее близок к потенциалу покоя клетки, последний имеет преимущественно калиевую природу. Это обусловлено тем, что высокая внутриклеточная концентрация К + не может существенно уменьшиться, так как ионы К + должны уравновешивать внутри клетки объемный отрицательный заряд анионов. Внутриклеточные анионы представляют собой в основном крупные органические молекулы (белки, остатки органических кислот ит.п.), которые не могут пройти через каналы в клеточной мембране. Концентрация этих анионов в клетке практически постоянна и их суммарный отрицательный заряд препятствует значительному выходу калия из клетки, поддерживая вместе с Na-K-насосом его высокую внутриклеточную концентрацию . Однако основная роль в первоначальном установлении внутри клетки высокой концентрации ионов калия и низкой концентрации ионов натрия принадлежит Na-K-насосу.
Распределение ионов С1 устанавливается в соответствии с мембранным потенциалом, поскольку в клетке нет специальных механизмов поддержания концентрации СГ. Поэтому вследствие отрицательного заряда хлора его распределение оказывается обратным по отношению к распределению калия на мембране (см. табл. 11.1). Таким образом, концентрационные диффузии К + из клетки и С1 в клетку практически уравновешиваются мембранным потенциалом покоя клетки.
Что касается Na + , то в покое его диффузия направлена в клетку под действием как градиента концентрации, так и электрического поля мембраны и вход Na + в клетку ограничивается в покое только малой проницаемостью мембраны для натрия (закрыты натриевые каналы). Действительно, Ходжкин и Катц экспериментально установили, что в состоянии покоя проницаемости мембраны аксона кальмара для К + , Na + и СГ относятся как 1: 0,04: 0,45. Таким образом, в состоянии покоя клеточная мембрана малопроницаема только для Na + , а для СГ она проницаема почти так же хорошо, как и для К + . В нервных клетках проницаемость для СГ обычно ниже, чем для К + , но в мышечных волокнах проницаемость для СГ даже несколько преобладает.
Несмотря на малую проницаемость клеточной мембраны для Na + в покое, существует, хотя и весьма малый, пассивный перенос Na + в клетку. Этот ток Na + должен был бы приводить к снижению разности потенциалов на мембране и к выходу К + из клетки, что вело бы в конечном итоге к выравниванию концентраций Na + и К + по обе стороны мембраны. Этого не происходит благодаря работе Na + - К + -насоса, компенсирующего токи утечки Na + и К + и поддерживающего таким образом нормальные значения внутриклеточных концентраций этих ионов и, следовательно, нормальную величину потенциала покоя клетки.
Для большинства клеток мембранный потенциал покоя составляет (-бО)-(-ЮО) мВ. На первый взгляд может показаться, что это малая величина, но надо учесть, что толщина мембраны тоже мала (8-10 нм), так что напряженность электрического поля в клеточной мембране огромна и составляет около 10 млн вольт на 1 м (или 100 кВ на 1 см):
Воздух, например, не выдерживает такой напряженности электрического поля (электрический пробой в воздухе наступает при 30 кВ/см), а мембрана выдерживает. Это нормальное условие ее деятельности, поскольку именно такое электрическое поле необходимо для поддержания разности концентраций ионов натрия, калия и хлора на мембране.
Величина потенциала покоя, различная у клеток, может изменяться при изменении условий их жизнедеятельности. Так, нарушение биоэнергетических процессов в клетке, сопровождающееся падением внутриклеточного уровня макро- эргических соединений (в частности, АТФ), прежде всего исключает компоненту потенциала покоя, связанную с работой Ма + -К + -АТФ-азы.
Повреждение клетки приводит обычно к повышению проницаемости клеточных мембран, в результате чего различия в проницаемости мембраны для ионов калия и натрия уменьшаются; потенциал покоя при этом уменьшается, что может вызвать нарушение ряда функций клетки, например возбудимости.
- Поскольку внутриклеточная концентрация калия поддерживается почти постоянной, то даже относительно небольшие изменения внеклеточной концентрации К* могут оказывать заметное влияние на потенциалпокоя и на деятельность клетки. Подобные изменения концентрации К"в плазме крови происходят при некоторых патологиях (например, припочечной недостаточности).

Чтобы провести сигнал от предшествующей клетки до последующей, нейрон генерирует электрические сигналы внутри себя. Твои движения глазами при чтении этого абзаца, ощущение мягкого кресла под попой, восприятие музыки из наушников и многое другое основаны на том, что внутри тебя проходят сотни миллиардов электрических сигналов. Такой сигнал может зародиться в спинном мозге и пройти до кончика пальца ноги по длинному аксону. Или может преодолеть ничтожно малое расстояние в глубинах мозга, ограничиваясь пределами интернейрона с короткими отростками. Любой нейрон, получивший сигнал, прогоняет его через свое тело и выросты, и этот сигнал имеет электрическую природу.
Еще в 1859 году ученые смогли измерить скорость, с которой передаются эти электрические сигналы. Оказалось, что электричество, передаваемое по живому аксону, принципиально отличается от электрического тока в металлах. По металлическому проводу электрический сигнал передается со скоростью, близкой к скорости света (300 000 километров в секунду), ведь в металле много свободных электронов. Однако, несмотря на эту скорость, сигнал ощутимо ослабевает, преодолевая большие расстояния. Если бы по аксонам сигналы передавались тем же способом, которым передаются в металлах, то нервный импульс, идущий от нервного окончания в коже большого пальца твоей ноги, полностью затухал бы, не достигая твоего мозга — электрическое сопротивление органической материи слишком велико, а сигнал слишком слаб.
Исследования показали, что электричество передается по аксонам намного медленнее, чем по проводам, и что в основе этой передачи лежит неизвестный ранее механизм, в результате которого сигнал распространяется со скоростью около 30 метров в секунду. Электрические сигналы, идущие по нервам, в отличие от сигналов, идущих по проводам, не ослабевают по ходу своего движения. Причина этого в том, что нервные окончания не пропускают через себя сигнал пассивно, просто позволяя имеющимся в них заряженным частицам передавать его друг другу. Они являются в каждой своей точке активным излучателем этого сигнала, ретранслируя его, и подробное описание этого механизма потребует отдельной главы. Таким образом, пожертвовав высокой скоростью проведения нервных импульсов, за счет активной передачи сигнала нейрон получает гарантию того, что возникший в большом пальце ноги сигнал достигнет спинного мозга, нисколько не ослабев.

Чтобы наблюдать прохождение электрической волны возбуждения, или потенциала действия (action potential [‘ækʃən pə’tenʃəl] ), в живой клетке, достаточно простого устройства: один конец тонкой металлической проволоки помещается на наружную поверхность аксона сенсорного нейрона кожи, а другой подводится к самописцу, чертящему линию вверх при усилении сигнала, и вниз — при ослаблении. Каждое прикосновение к коже вызывает один или несколько потенциалов действия. При возникновении каждого потенциала самописец рисует узкий длинный пик.
Потенциал действия сенсорного нейрона длится всего лишь около 0,001 секунды и включает две фазы: быстрого нарастания, достигающего пика, а затем почти столь же быстрого спада возбуждения, приводящего к исходному положению. И тут самописец сообщает неожиданный факт: все потенциалы действия, возникающие в одной и той же нервной клетке, примерно одинаковы. Это можно увидеть на картинке слева: все пики, нарисованные самописцем, имеют примерно одну и ту же форму и амплитуду независимо от того, насколько сильным или продолжительным было прикосновение к коже, их вызвавшее. Слабое поглаживание или ощутимый щипок будут переданы потенциалами действия одной и той же величины. Потенциал действия представляет собой постоянный сигнал, подчиняющийся принципу «все или ничего»: после превышения раздражителем некоего порогового значения возникает всегда примерно одинаковый сигнал, не больше и не меньше обычного. А если раздражитель меньше порогового значения, то сигнал вовсе не будет передаваться: например, можно так легко коснуться кожи кончиком пера, что это прикосновение не будет чувствоваться.
Принцип «все или ничего» в возникновении потенциала действия вызывает новые вопросы. Как сенсорный нейрон сообщает о силе раздражителя - сильное или слабое давление, яркий или тусклый свет? Как он сообщает о продолжительности действия раздражителя? Наконец, как нейроны отличают один тип сенсорной информации от другого - например, как они отличают прикосновение от боли, света, запаха или звука? И как они отличают сенсорную информацию для восприятия от моторной информации для действия?

Эволюция решила вопрос о том, как сообщить о силе раздражителя, с помощью использования одного и того же вида сигналов одной и той же величины: эта сила определяется частотой (frequency [‘friːkwənsɪ] ), с которой испускаются потенциалы действия. Слабый раздражитель, например легкое прикосновение к руке, приводит к испусканию всего двух-трех потенциалов действия в секунду, в то время как сильное давление, как при щипке или ударе по локтю, может вызвать очередь из сотни потенциалов действия в секунду. При этом продолжительность ощущения определяется продолжительностью возникновения потенциалов действия.
Используют ли нейроны разные электрические коды, сообщая мозгу, что несут информацию о разных раздражителях, таких как боль, свет или звук? Оказалось, что нет! Это удивительно, но между потенциалами действия, генерируемыми нейронами из различных сенсорных систем (например, зрительной или тактильной), разница весьма незначительна! Таким образом, характер и природа ощущения не зависят от различий в потенциалах действия (что открывает довольно захватывающую перспективу для размышлений на тему «матрицы» из одноименного фильма). Нейрон, передающий слуховую информацию, устроен точно так же, как нейрон из зрительной нервной цепи, и проводят они одни и те же потенциалы действия, одним и тем же способом. Без знания, к какой нервной цепи принадлежит конкретный нейрон, только по анализу его функционирования невозможно определить, какую информацию он несет.
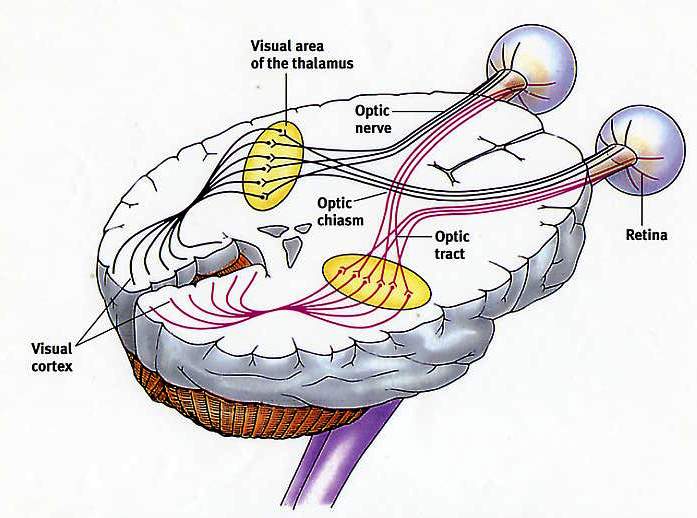
Природа передаваемой информации зависит прежде всего от типа возбуждаемых нервных волокон и специфических систем мозга, с которыми эти волокна связаны. Ощущения каждого типа передаются по своим проводящим путям, и разновидность передаваемой нейроном информации зависит именно от пути, в состав которого входит этот нейрон. В любом сенсорном проводящем пути информация передается от первого сенсорного нейрона (рецептора, реагирующего на внешний раздражитель, например прикосновение, запах или свет) к специализированным нейронам в спинном или головном мозге. Таким образом, зрительная информация отличается от слуховой лишь тем, что передается по другим проводящим путям, начинающимся в сетчатке глаза и заканчивающимся в участке мозга, который отвечает за визуальное восприятие.
Сигналы, посылаемые от моторных нейронов мозга к мышцам, также почти идентичны передаваемым по сенсорным нейронам от кожи в мозг. Они подчиняются тому же принципу «все или ничего», так же передают интенсивность сигнала с помощью частоты потенциалов действия, и так же результат сигнала зависит только от того, в какую нервную цепь включен этот нейрон. Таким образом, быстрая череда потенциалов действия, идущая по определенному проводящему пути, вызывает именно движение твоих пальцев, а не, скажем, восприятие разноцветных огней, лишь потому, что данный путь связан с мышцами рук, а не с сетчаткой глаз.

Универсальность потенциалов действия не ограничивается схожестью их проявления в разных нейронах, находящихся в пределах одного организма. Они настолько одинаковы у разных животных, что даже умудренный опытом исследователь не способен точно отличить запись потенциала действия нервного волокна кита, мыши, обезьяны или его научного руководителя. Тем не менее потенциалы действия в разных клетках не являются идентичными: небольшая разница в их амплитуде и длительности все же есть, и утверждение «все потенциалы действия одинаковы» так же неточно, как и «все бугенвиллии одинаковы».
Итак, каждый нейрон передает сигнал через свое тело и отростки одним и тем же образом. Все разнообразие информации, получаемой нами от сенсорных нейронов, все движения, которые может совершать наше тело — результат передачи единственного типа сигналов внутри нейронов. Осталась «мелочь»: понять, что же это за сигнал и как он передается.

Мы привычно отделяем все, что считаем живой природой, в том числе и себя самих, от «неживых» вещей, в том числе металлов и передающегося через них электрического тока. Тем удивительнее осознавать, что в наших телах металлы не просто присутствуют — они необходимы, без них тело не сможет существовать. Электрический ток — явление не разовое, а непрерывно возникающее в сотне миллиардов нейронов, пронизавших своими отростками все наше тело. Прямо сейчас ты можешь ощутить самые разные признаки его присутствия: то, что ты осознаешь этот текст, есть результат бесчисленных передач электрического тока. Чувство голода и удовольствие от запаха готовящейся еды, само восприятие этого запаха, прикосновение залетевшего в окно ветра к твоей коже… Перечислять можно бесконечно. И желание понять, каким же образом все это происходит, также складывается из возникающих в нейронах электрических импульсов.
Так как целью этой главы является сообщение лишь самой общей информации о прохождении нервного импульса, то здесь же необходимо рассмотреть ту среду, в которой он возникает, те условия в клетке, которые делают возможным его возникновение и передачу. Поэтому стоит начать с изучения плацдарма, на котором будут развиваться события, а именно с нейрона в состоянии покоя (dormant state [‘dɔːmənt steɪt] ).

Еще в середине прошлого века ученые нашли способ установить, в какой части нейрона существует электрический заряд. Для этого используют вольтметр (voltmeter [‘vəultˌmiːtə] ) (прибор для измерения напряжения электрического поля) с двумя электродами. Один электрод помещают внутрь нейрона, располагая его близко к клеточной мембране, а второй электрод находится в окружающей нейрон среде, с другой стороны той же мембраны. Вольтметр показывает, что с разных сторон клеточной мембраны существуют электрические заряды , отрицательный внутри клетки и положительный снаружи. Существование таких разнополюсных электрических зарядов по обе стороны мембраны создает электрическое поле, важной характеристикой которого является потенциал . Потенциал, говоря простым языком, это способность совершать работу, например работу по перетаскиванию заряженной частицы с места на место. Чем больше отрицательных зарядов накопилось по одну сторону, и чем больше положительных — по другую сторону мембраны, тем сильнее создаваемое ими электрическое поле, и тем с большей силой они способны перетаскивать туда-сюда заряженные частицы. Разницу между внешним и внутренним электрическими зарядами называют мембранным потенциалом (membrane potential [‘membreɪn pə’tenʃəl] ) покоя. Для нейрона он равен примерно 70 мВ (милливольт), то есть 70 тысячных вольта или семь сотых вольта. Для сравнения, разность потенциалов в батарейке АА равна 1,5 вольта — в 20 раз больше. То есть мембранный потенциал покоя нейрона всего лишь в 20 раз слабее, чем между клеммами батарейки АА — довольно большой, получается. Электрический потенциал существует только на мембране, и в других своих частях нейрон электрически нейтрален.
Если написать более точно, то мембранный потенциал покоя нейрона равен -70 мВ (минус семьдесят милливольт). Знак минус означает лишь то, что отрицательный заряд находится именно внутри клетки, а не снаружи, и таким образом создаваемое электрическое поле способно перетаскивать через мембрану внутрь клетки положительно заряженные ионы.
Действующие лица в создании мембранного потенциала покоя:

1 . В клеточной мембране нейрона существуют каналы, по которым несущие электрический заряд ионы могут путешествовать сквозь нее. При этом мембрана не является всего лишь пассивной «перегородкой» между внутренней средой нейрона и окружающей его межклеточной жидкостью: специальные белки, внедренные в плоть мембраны, открывают и закрывают эти каналы, и таким образом мембрана контролирует прохождение ионов — атомов, имеющих электрический заряд. Накапливая отрицательно заряженные ионы внутри клетки, нейрон увеличивает количество отрицательных зарядов внутри, тем самым приводя к увеличению положительных зарядов снаружи, и таким образом усиливается электрический потенциал. Так как протон имеет положительный заряд, а электрон отрицательный, то при избытке протонов получается положительно заряженный ион, а при избытке электронов — отрицательно заряженный. Если хочется более подробной информации об атомах и ионах, можно вернуться в . Важно понимать, что мембранный потенциал существует именно на границе клеточной мембраны, а жидкости в целом внутри и вне нейрона остаются электрически нейтральными. Ионы, для которых мембрана проницаема, остаются вблизи нее, поскольку положительные и отрицательные заряды взаимно притягиваются друг к другу. В результате снаружи мембраны образуется слой «сидящих» на ней положительных ионов, а внутри — отрицательных. Таким образом, мембрана играет роль электрической емкости, разделяющей заряды, внутри которой есть электрическое поле. Мембрана, поэтому, является природным конденсатором.
2 . отрицательно заряженные протеины , находящиеся внутри нейрона возле внутренней поверхности мембраны. Заряд протеинов всегда остается одним и тем же и является только частью общего заряда внутренней поверхности мембраны. В отличие от ионов, протеины не могут выходить из клетки и заходить в нее — для этого они слишком большие. Общий заряд меняется в зависимости от количества находящихся возле мембраны положительно заряженных ионов, концентрация которых может меняться за счет их перехода из клетки наружу, и извне вовнутрь.
3 . положительно заряженные ионы калия (К +) могут свободно перемещаться между внутренней и внешней средой, когда нейрон находится в состоянии покоя. Перемещаются они через постоянно открытые проточные калиевые каналы (flow potassium passage ), через которые могут пройти только ионы К + , и ничто другое. Проточными называются каналы, не имеющие ворот, а значит открытые при любом состоянии нейрона. Внутри клетки ионов калия гораздо больше, чем снаружи. Это происходит за счет постоянной работы натрий-калиевого насоса (про него будет рассказано ниже), поэтому в состоянии покоя нейрона ионы К + начинают перемещаться во внешнюю среду, поскольку концентрация одного и того же вещества стремится выровняться в общей системе. Если мы в бассейн с водой в одном углу выльем какое-то вещество, то его концентрация в этом углу будет очень большой, а в других частях бассейна — нулевой или очень маленькой. Однако, спустя некоторое время мы обнаружим, что концентрация этого вещества выровнялась по всему бассейну за счет броуновского движения. В этом случае говорят о «парциальном давлении» того или иного вещества, будь это жидкость или газ. Если в одном углу бассейна будет вылит спирт, то образуется большая разница в концентрации спирта между этим углом и остальным бассейном. Возникнет парциальное давление молекул спирта, и они постепенно распределятся равномерно по бассейну так, что парциальное давление исчезнет, поскольку концентрация молекул спирта везде выровняется. Таким образом, ионы К + уносят с собой положительный заряд из нейрона, уходя наружу за счет парциального давления, которое сильнее, чем сила притяжения отрицательно заряженных протеинов, в том случае, если разница в концентрации ионов внутри и снаружи клетки достаточно велика. Так как внутри остаются отрицательно заряженные протеины, то таким образом на внутренней стороне мембраны формируется отрицательный заряд. Для ясного понимания работы клеточных механизмов важно помнить, что несмотря на постоянное вытекание ионов калия из клетки, внутри нейрона их всегда больше, чем снаружи.
4 . положительно заряженные ионы натрия (Na +) находятся с внешней стороны мембраны и создают там положительный заряд. Во время фазы покоя нейрона натриевые каналы клетки закрыты , и Na + не могут пройти внутрь, а их концентрация снаружи повышается за счет работы натрий-калиевого насоса, выводящего их из нейрона.
5 . роль отрицательно заряженных ионов хлора (Cl —) и положительно заряженных ионов кальция (Ca 2+) для создания мембранного потенциала невелика, поэтому их поведение пока останется за кадром.
Формирование мембранного потенциала покоя проходит в два этапа:
Этап I . создается небольшая (-10 мВ) разница потенциалов с помощью натрий-калиевого насоса .

В отличие от других каналов мембраны, натрий-калиевый канал способен пропускать через себя и ионы натрия, и ионы калия. Причем Na + может пройти сквозь него только из клетки наружу, а К + снаружи внутрь. Один цикл работы этого канала включается в себя 4 шага:
1 . «ворота» натрий-калиевого канала открыты только с внутренней стороны мембраны, и туда заходят 3 Na +

2 . присутствие Na + внутри канала воздействует на него так, что он может частично разрушить одну молекулу АТФ (ATP ) (аденозинтрифосфата ), (adenosine triphosphate [ə’dɛnəsiːn trai’fɔsfeɪt] ) являющуюся «аккумулятором» клетки, запасающим энергию и отдающим ее при необходимости. При таком частичном разрушении, заключающемся в отщеплении от конца молекулы одной фосфатной группы PO 4 3− , выделяется энергия, которая как раз и расходуется на перенос Na + во внешнее пространство.
3 . когда канал открывается для того, чтобы Na + вышел наружу, он остается открытым, и в него попадают два иона К + — их притягивают отрицательные заряды протеинов изнутри. То, что в канале, вмещающем три иона натрия, помещается всего два иона калия, вполне логично: атом калия имеет больший диаметр.
4 . присутствие ионов калия теперь в свою очередь воздействует на канал так, что внешние «ворота» закрываются, а внутренние открываются, и К + поступают во внутреннюю среду нейрона.
Таким образом работает натрий-калиевый насос, «обменивая» три иона натрия на два иона калия. Так как электрический заряд у Na + и К + одинаковый, получается что из клетки выводится три положительных заряда, а внутрь попадает только два. За счет этого внутренний положительный заряд клеточной мембраны снижается, а внешний — увеличивается. К тому же создается разница в концентрации Na + и К + по разные стороны мембраны:
=) снаружи клетки оказывается много ионов натрия, а внутри — мало. При этом натриевые каналы закрыты, и попасть назад в клетку Na + не может, и далеко от мембраны он не уходит, так как притягивается существующим с внутренней стороны мембраны отрицательным зарядом.
=) внутри клетки много ионов калия, а вот снаружи их мало, и это приводит к вытеканию К + из клетки через открытые во время фазы покоя нейрона калиевые каналы.

Этап II формирования мембранного потенциала покоя как раз основан на этом вытекании ионов калия из нейрона. На рисунке слева показан ионный состав мембраны в начале второго этапа формирования потенциала покоя: множество К + и отрицательно заряженных протеинов (обозначенных А 4-) внутри, и облепившие мембрану снаружи Na + . Перемещаясь во внешнюю среду, ионы калия уносят из клетки свои положительные заряды, при этом суммарный заряд внутренней мембраны снижается. Так же как положительные ионы натрия, вытекшие из клетки ионы калия остаются снаружи мембраны, притягиваемые внутренним отрицательным зарядом, и внешний положительный заряд мембраны складывается из суммы зарядов Na + и К + . Несмотря на вытекание через проточные каналы, внутри клетки ионов калия всегда больше, чем снаружи.
Возникает вопрос: почему ионы калия не продолжают вытекать наружу до того момента, пока их количество внутри клетки и вне ее не станет одинаковым, то есть до тех пор, пока не исчезнет парциальное давление, создаваемое этими ионами? Причина этого заключается в том, что когда К + покидают клетку, снаружи увеличивается положительный заряд, а внутри образуется избыток отрицательного заряда. Это снижает желание ионов калия выходить из клетки, ведь наружный положительный заряд их отталкивает, а внутренний отрицательный притягивает. Поэтому через какое-то время К + перестают вытекать несмотря на то, что во внешней среде их концентрация ниже, чем во внутренней: влияние зарядов по разные стороны мембраны превышает силу парциального давления, то есть превышает стремление К + распределиться равномерно в жидкости внутри и вне нейрона. В момент достижения этого равновесия мембранный потенциал нейрона и останавливается примерно на -70 мВ.
Как только нейроном достигнут мембранный потенциал покоя, он готов для возникновения и проведения потенциала действия, про который речь пойдет в следующей цитологической главе.
Таким образом, подытожим : неравномерность распределения ионов калия и натрия по обе стороны мембраны вызвана действием двух соперничающих сил: а) силой электрического притяжения и отталкивания, и б) силой парциального давления, возникающего при разнице в концентрациях. Работа этих двух соперничающих сил протекает в условиях существования по-разному устроенных натриевых, калиевых и натриево-калиевого каналов, которые выступают в роли регуляторов действия этих сил. Калиевый канал является проточным, то есть он всегда открыт в состоянии покоя нейрона, так что ионы К + могут спокойно ходить туда-сюда под воздействием сил электрического отталкивания/притяжения и под воздействием силы, вызванные парциальным давлением, то есть разницей в концентрации этих ионов. Натриевый канал всегда закрыт в состоянии покоя нейрона, так что через них ионы Na + ходить не могут. И, наконец, натриево-калиевый канал, устроенный так, что он работает как насос, который при каждом цикле выгоняет три иона натрия наружу, и загоняет два иона калия внутрь.
Вся эта конструкция и обеспечивает возникновение мембранного потенциала покоя нейрона: т.е. состояния, при котором достигается две вещи:
а) внутри есть отрицательный заряд, а снаружи — положительный.
б) внутри много ионов К + , облепивших отрицательно заряженные части протеинов, и таким образом возникает калиевое парциальное давление — стремление ионов калия выйти наружу для выравнивания концентрации.
в) снаружи много ионов Na + , образующих отчасти пары с ионами Cl — . И таким образом возникает натриевое парциальное давление — стремление ионов натрия войти внутрь клетки для выравнивания концентрации.
В результате работы калиево-натриевого насоса мы получаем три силы, существующие на мембране: силу электрического поля и силу двух парциальных давлений. Эти силы и начинают работать, когда нейрон выходит из состояния покоя.